

Meiotic sister chromatid exchanges are rare in C. elegans
- PMID: 33740426
- PMCID: PMC8051885
- DOI: 10.1016/j.cub.2020.11.018
Meiotic sister chromatid exchanges are rare in C. elegans
Abstract
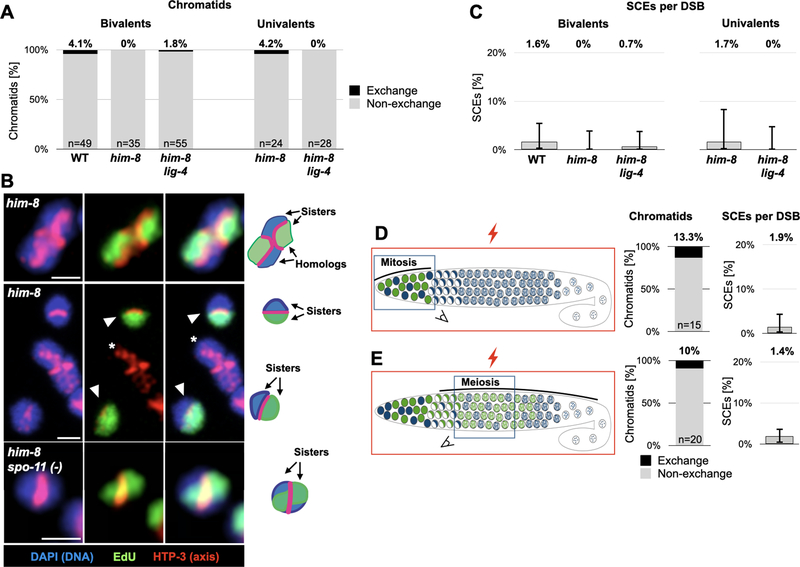
Sexual reproduction shuffles the parental genomes to generate new genetic combinations. To achieve that, the genome is subjected to numerous double-strand breaks, the repair of which involves two crucial decisions: repair pathway and repair template.1 Use of crossover pathways with the homologous chromosome as template exchanges genetic information and directs chromosome segregation. Crossover repair, however, can compromise the integrity of the repair template and is therefore tightly regulated. The extent to which crossover pathways are used during sister-directed repair is unclear because the identical sister chromatids are difficult to distinguish. Nonetheless, indirect assays have led to the suggestion that inter-sister crossovers, or sister chromatid exchanges (SCEs), are quite common.2-11 Here we devised a technique to directly score physiological SCEs in the C. elegans germline using selective sister chromatid labeling with the thymidine analog 5-ethynyl-2'-deoxyuridine (EdU). Surprisingly, we find SCEs to be rare in meiosis, accounting for <2% of repair events. SCEs remain rare even when the homologous chromosome is unavailable, indicating that almost all sister-directed repair is channeled into noncrossover pathways. We identify two mechanisms that limit SCEs. First, SCEs are elevated in the absence of the RecQ helicase BLMHIM-6. Second, the synaptonemal complex-a conserved interface that promotes crossover repair12,13-promotes SCEs when localized between the sisters. Our data suggest that crossover pathways in C. elegans are only used to generate the single necessary link between the homologous chromosomes. Noncrossover pathways repair almost all other breaks, regardless of the repair template.
Keywords: BLM; C. elegans; DNA repair; EdU; RecQ helicase; SCE; crossover; meiosis; synaptonemal complex.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



Comment in
-
Meiosis: The elusive sister chromatid repair.Curr Biol. 2021 May 10;31(9):R454-R456. doi: 10.1016/j.cub.2021.03.093. Curr Biol. 2021. PMID: 33974876
References
-
- Schwacha A, and Kleckner N (1994). Identification of joint molecules that form frequently between homologs but rarely between sister chromatids during yeast meiosis. Cell 76, 51–63. - PubMed
-
- McKim KS, Green-Marroquin BL, Sekelsky JJ, Chin G, Steinberg C, Khodosh R, and Hawley RS (1998). Meiotic synapsis in the absence of recombination. Science 279, 876–878. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
