

Meiotic DNA break repair can utilize homolog-independent chromatid templates in C. elegans
- PMID: 33740427
- PMCID: PMC8189575
- DOI: 10.1016/j.cub.2021.03.008
Meiotic DNA break repair can utilize homolog-independent chromatid templates in C. elegans
Abstract
During meiosis, the maintenance of genome integrity is critical for generating viable haploid gametes.1 In meiotic prophase I, double-strand DNA breaks (DSBs) are induced and a subset of these DSBs are repaired as interhomolog crossovers to ensure proper chromosome segregation. DSBs not resolved as crossovers with the homolog must be repaired by other pathways to ensure genome integrity.2 To determine if alternative repair templates can be engaged for meiotic DSB repair during oogenesis, we developed an assay to detect sister and/or intra-chromatid repair events at a defined DSB site during Caenorhabditis elegans meiosis. Using this assay, we directly demonstrate that the sister chromatid or the same DNA molecule can be engaged as a meiotic repair template for both crossover and noncrossover recombination, with noncrossover events being the predominant recombination outcome. We additionally find that the sister or intra-chromatid substrate is available as a recombination partner for DSBs induced throughout meiotic prophase I, including late prophase when the homolog is unavailable. Analysis of noncrossover conversion tract sequences reveals that DSBs are processed similarly throughout prophase I. We further present data indicating that the XPF-1 nuclease functions in late prophase to promote sister or intra-chromatid repair at steps of recombination following joint molecule processing. Despite its function in sister or intra-chromatid repair, we find that xpf-1 mutants do not exhibit severe defects in progeny viability following exposure to ionizing radiation. Overall, we propose that C. elegans XPF-1 may assist as an intersister or intrachromatid resolvase only in late prophase I.
Keywords: C. elegans; DNA repair; double-strand DNA break; genome integrity; germ line; meiosis; oogenesis; recombination; sister chromatid; worms.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare that they have no competing interests.
Figures
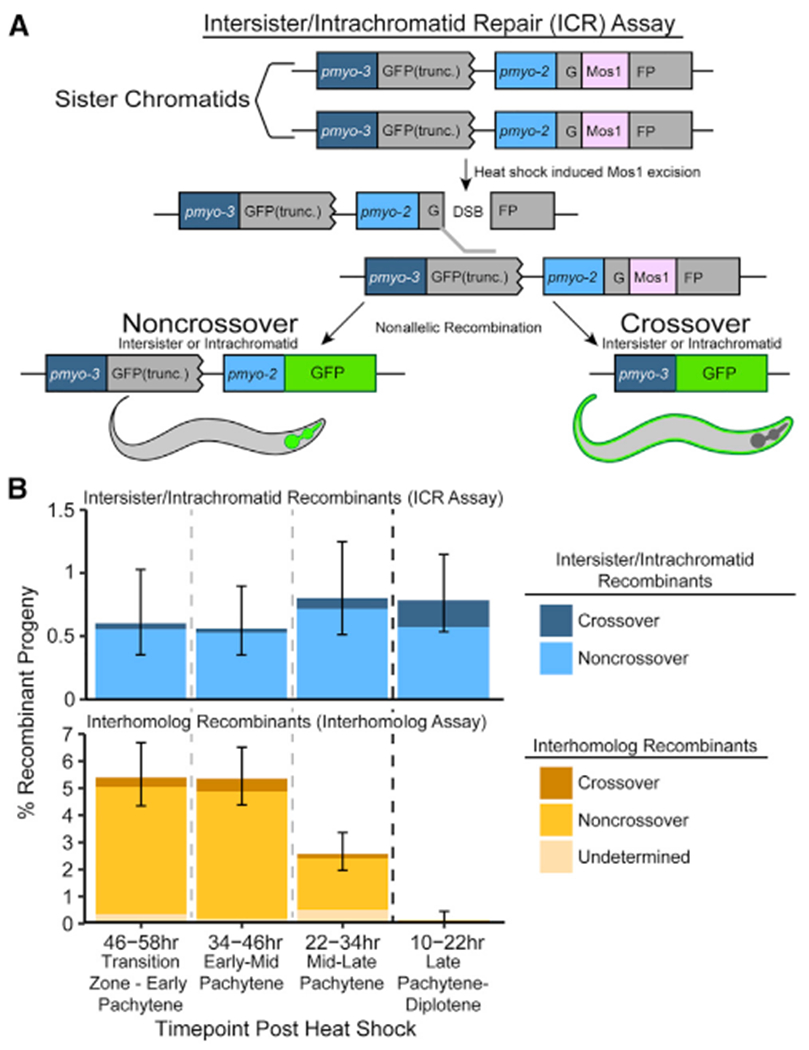
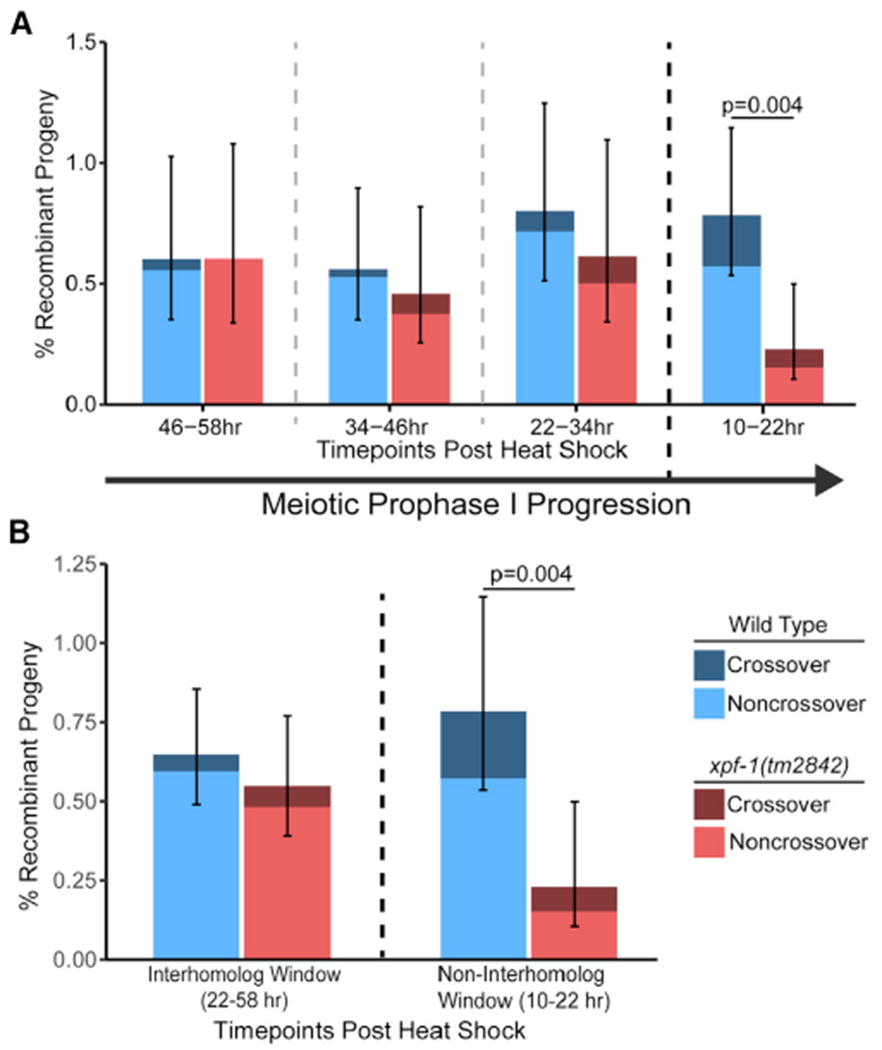
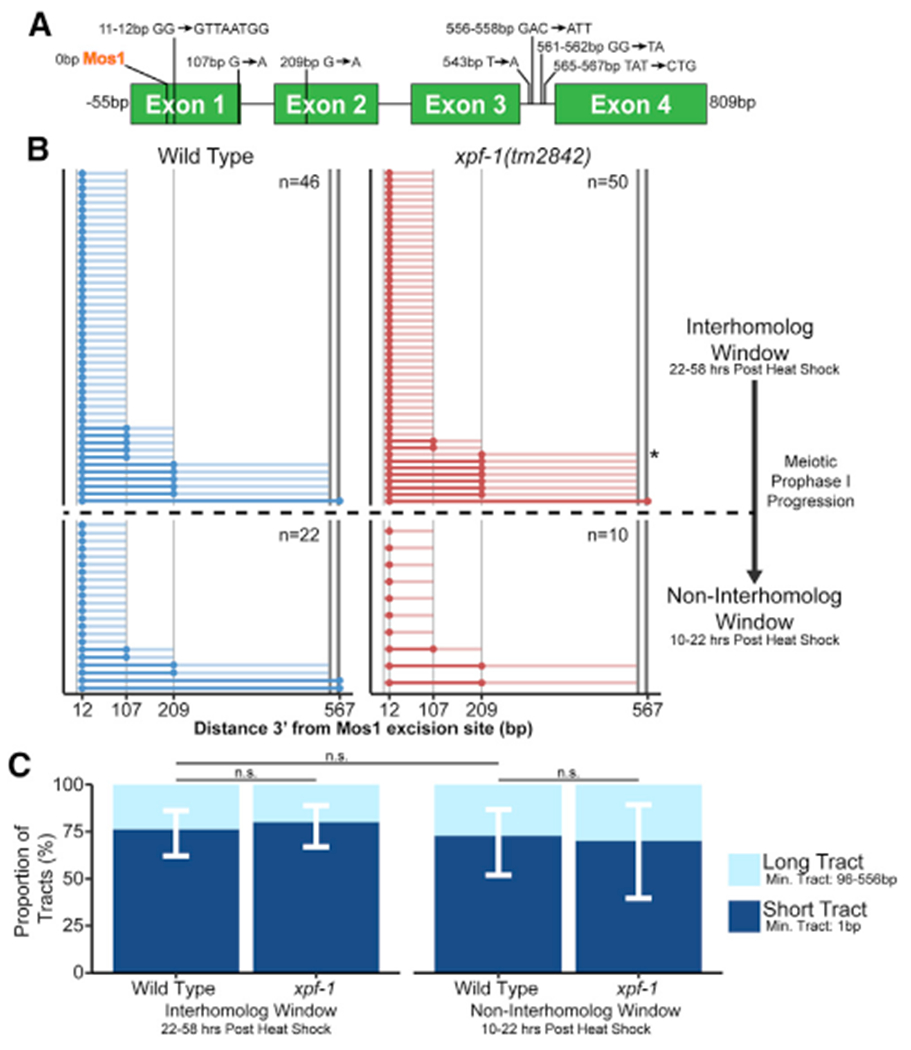
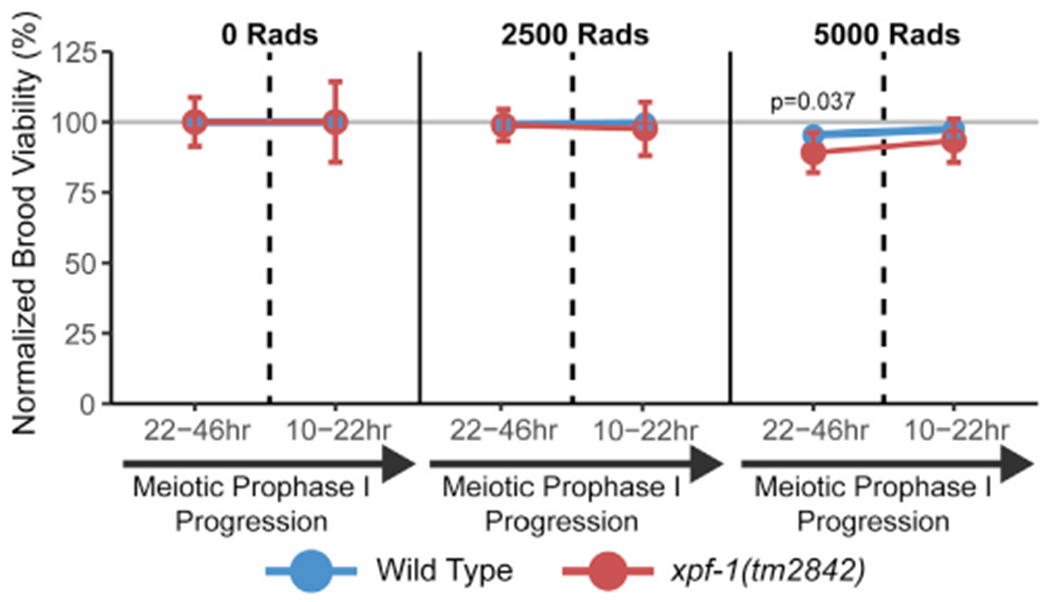
Comment in
-
Meiosis: The elusive sister chromatid repair.Curr Biol. 2021 May 10;31(9):R454-R456. doi: 10.1016/j.cub.2021.03.093. Curr Biol. 2021. PMID: 33974876
References
-
- Handel MA, and Schimenti JC (2010). Genetics of mammalian meiosis: regulation, dynamics and impact on fertility. Nat. Rev. Genet. 11,124–136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
