

Functional Analysis of Phenazine Biosynthesis Genes in Burkholderia spp
- PMID: 33741619
- PMCID: PMC8208154
- DOI: 10.1128/AEM.02348-20
Functional Analysis of Phenazine Biosynthesis Genes in Burkholderia spp
Abstract
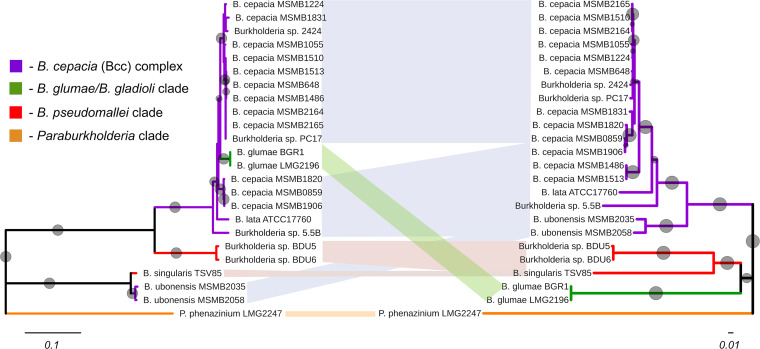
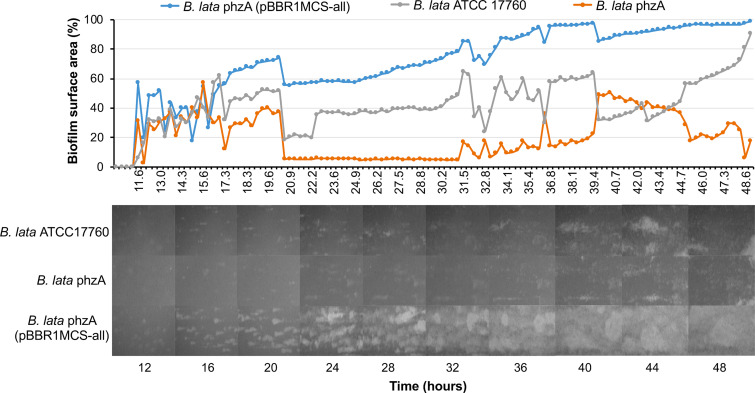
Burkholderia encompasses a group of ubiquitous Gram-negative bacteria that includes numerous saprophytes as well as species that cause infections in animals, immunocompromised patients, and plants. Some species of Burkholderia produce colored, redox-active secondary metabolites called phenazines. Phenazines contribute to competitiveness, biofilm formation, and virulence in the opportunistic pathogen Pseudomonas aeruginosa, but knowledge of their diversity, biosynthesis, and biological functions in Burkholderia is lacking. In this study, we screened publicly accessible genome sequence databases and identified phenazine biosynthesis genes in multiple strains of the Burkholderia cepacia complex, some isolates of the B. pseudomallei clade, and the plant pathogen B. glumae We then focused on B. lata ATCC 17760 to reveal the organization and function of genes involved in the production of dimethyl 4,9-dihydroxy-1,6-phenazinedicarboxylate. Using a combination of isogenic mutants and plasmids carrying different segments of the phz locus, we characterized three novel genes involved in the modification of the phenazine tricycle. Our functional studies revealed a connection between the presence and amount of phenazines and the dynamics of biofilm growth in flow cell and static experimental systems but at the same time failed to link the production of phenazines with the capacity of Burkholderia to kill fruit flies and rot onions.IMPORTANCE Although the production of phenazines in Burkholderia was first reported almost 70 years ago, the role these metabolites play in the biology of these economically important microorganisms remains poorly understood. Our results revealed that the phenazine biosynthetic pathway in Burkholderia has a complex evolutionary history, which likely involved horizontal gene transfers among several distantly related groups of organisms. The contribution of phenazines to the formation of biofilms suggests that Burkholderia, like fluorescent pseudomonads, may benefit from the unique redox-cycling properties of these versatile secondary metabolites.
Keywords: Burkholderia; biosynthesis; phenazine.
Copyright © 2021 American Society for Microbiology.
Figures





Similar articles
-
Quorum sensing and phenazines are involved in biofilm formation by Pseudomonas chlororaphis (aureofaciens) strain 30-84.Microb Ecol. 2006 Aug;52(2):289-301. doi: 10.1007/s00248-006-9064-6. Epub 2006 Aug 4. Microb Ecol. 2006. PMID: 16897305
-
Pseudomonas aeruginosa PumA acts on an endogenous phenazine to promote self-resistance.Microbiology (Reading). 2018 May;164(5):790-800. doi: 10.1099/mic.0.000657. Epub 2018 Apr 9. Microbiology (Reading). 2018. PMID: 29629858 Free PMC article.
-
Diversity and evolution of the phenazine biosynthesis pathway.Appl Environ Microbiol. 2010 Feb;76(3):866-79. doi: 10.1128/AEM.02009-09. Epub 2009 Dec 11. Appl Environ Microbiol. 2010. PMID: 20008172 Free PMC article.
-
Recent insights into the diversity, frequency and ecological roles of phenazines in fluorescent Pseudomonas spp.Environ Microbiol. 2013 Mar;15(3):675-86. doi: 10.1111/j.1462-2920.2012.02846.x. Epub 2012 Aug 13. Environ Microbiol. 2013. PMID: 22882648 Review.
-
Phenazines in plant-beneficial Pseudomonas spp.: biosynthesis, regulation, function and genomics.Environ Microbiol. 2018 Nov;20(11):3905-3917. doi: 10.1111/1462-2920.14395. Epub 2018 Oct 15. Environ Microbiol. 2018. PMID: 30159978 Review.
Cited by
-
Identification of Key Factors for Anoxic Survival of B. cenocepacia H111.Int J Mol Sci. 2022 Apr 20;23(9):4560. doi: 10.3390/ijms23094560. Int J Mol Sci. 2022. PMID: 35562951 Free PMC article.
-
Screening of natural phenazine producers for electroactivity in bioelectrochemical systems.Microb Biotechnol. 2023 Mar;16(3):579-594. doi: 10.1111/1751-7915.14199. Epub 2022 Dec 26. Microb Biotechnol. 2023. PMID: 36571174 Free PMC article.
-
Pangenome Analysis Reveals Novel Contact-Dependent Growth Inhibition System and Phenazine Biosynthesis Operons in Proteus mirabilis BL95 That Are Located in An Integrative and Conjugative Element.Microorganisms. 2024 Jun 28;12(7):1321. doi: 10.3390/microorganisms12071321. Microorganisms. 2024. PMID: 39065090 Free PMC article.
-
Insights into group-specific pattern of secondary metabolite gene cluster in Burkholderia genus.Front Microbiol. 2024 Jan 16;14:1302236. doi: 10.3389/fmicb.2023.1302236. eCollection 2023. Front Microbiol. 2024. PMID: 38293557 Free PMC article. Review.
-
Compilation of the Antimicrobial Compounds Produced by Burkholderia Sensu Stricto.Molecules. 2023 Feb 8;28(4):1646. doi: 10.3390/molecules28041646. Molecules. 2023. PMID: 36838633 Free PMC article. Review.
References
-
- Woods SE, Sokol PA. 2006. The genus Burkholderia, p 848–860. In Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E (ed), The Prokaryotes, vol 5. Springer, New York.
-
- Yabuuchi E, Kosako Y, Oyaizu H, Yano I, Hotta H, Hashimoto Y, Ezaki T, Arakawa M. 1992. Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol Immunol 36:1251–1275. 10.1111/j.1348-0421.1992.tb02129.x. - DOI - PubMed
-
- Estrada-de Los Santos P, Rojas-Rojas FU, Tapia-García EY, Vásquez-Murrieta MS, Hirsch AM. 2016. To split or not to split: an opinion on dividing the genus Burkholderia. Ann Microbiol 66:1303–1314. 10.1007/s13213-015-1183-1. - DOI
-
- Sawana A, Adeolu M, Gupta RS. 2014. Molecular signatures and phylogenomic analysis of the genus Burkholderia: proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front Genet 5:429. 10.3389/fgene.2014.00429. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
