

Anopheles ecology, genetics and malaria transmission in northern Cambodia
- PMID: 33742030
- PMCID: PMC7979810
- DOI: 10.1038/s41598-021-85628-1
Anopheles ecology, genetics and malaria transmission in northern Cambodia
Erratum in
-
Publisher Correction: Anopheles ecology, genetics and malaria transmission in northern Cambodia.Sci Rep. 2021 Jun 9;11(1):12553. doi: 10.1038/s41598-021-91552-1. Sci Rep. 2021. PMID: 34108534 Free PMC article. No abstract available.
Abstract
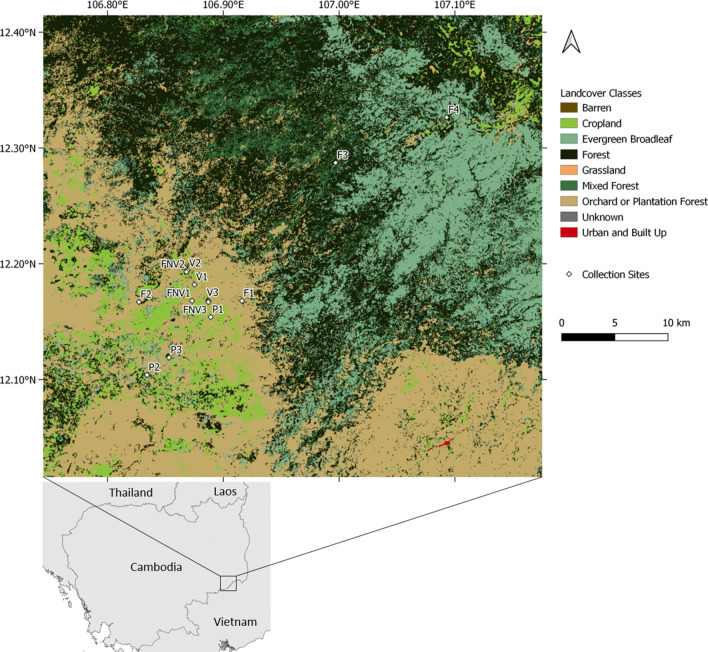
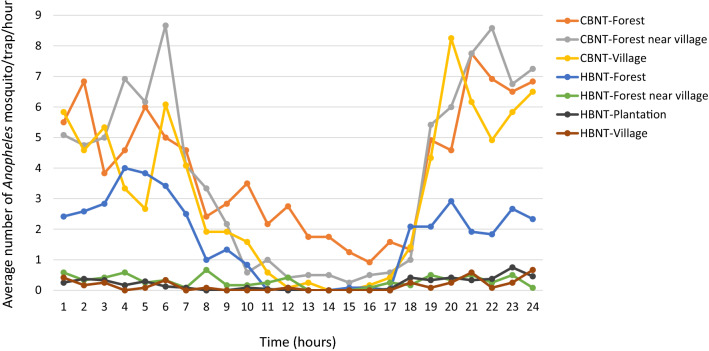
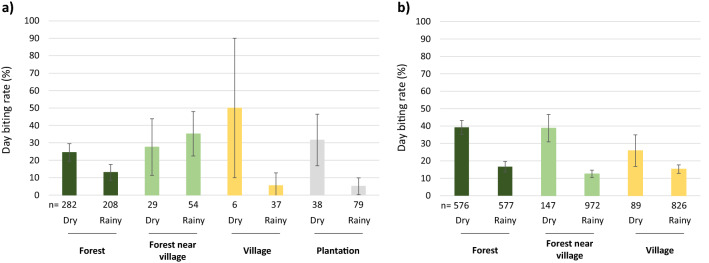
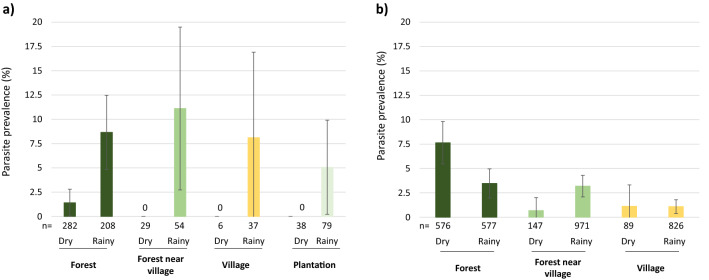
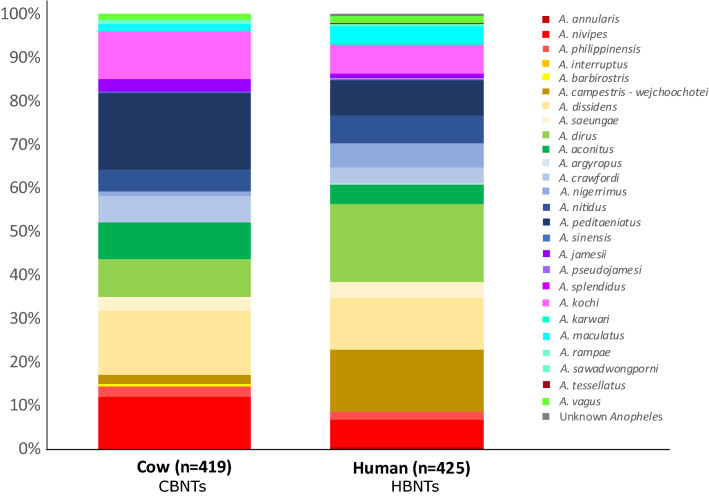
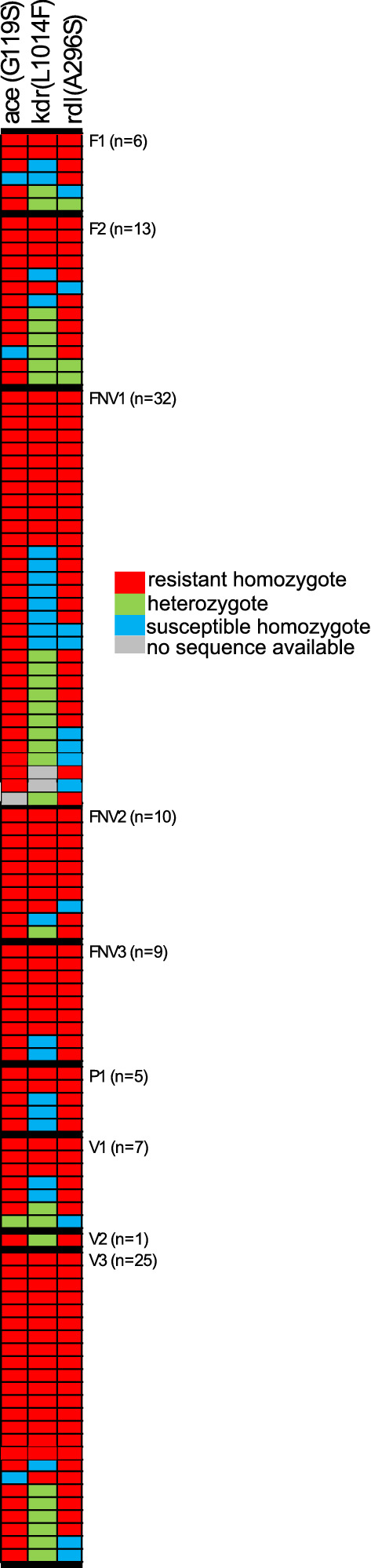
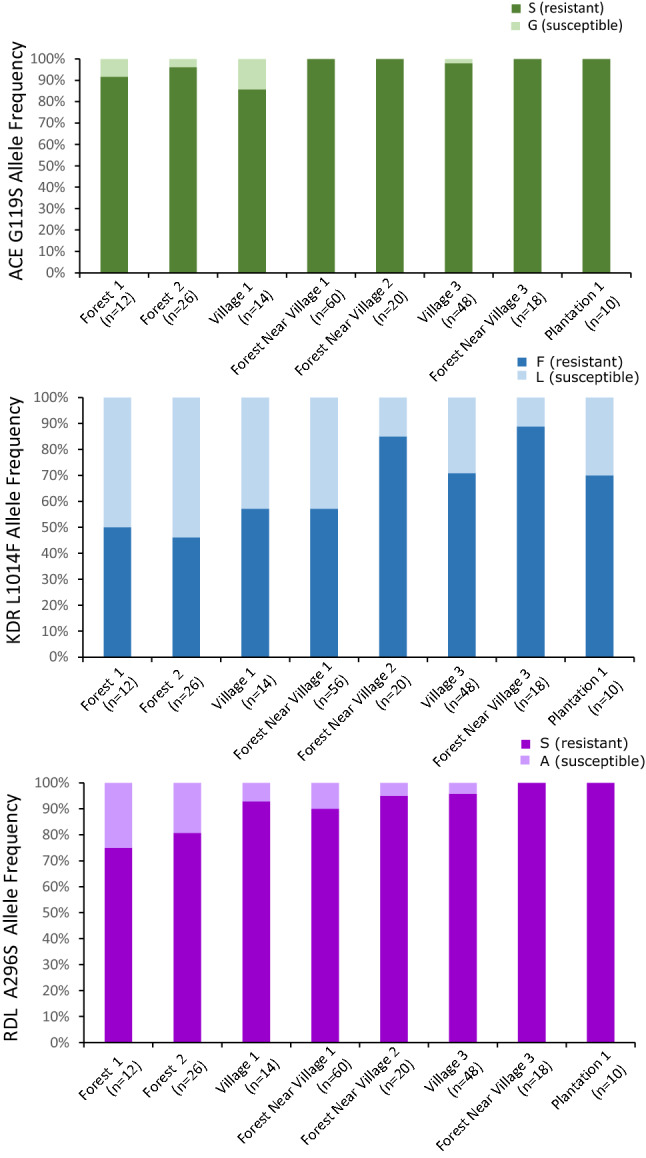
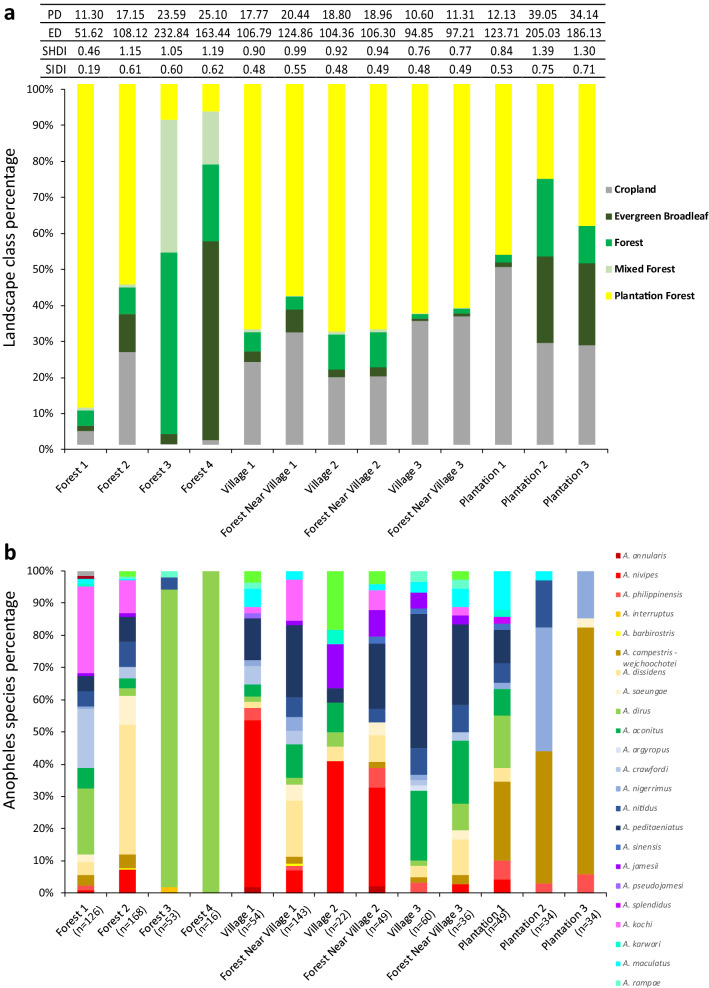
In the Greater Mekong Subregion, malaria cases have significantly decreased but little is known about the vectors or mechanisms responsible for residual malaria transmission. We analysed a total of 3920 Anopheles mosquitoes collected during the rainy and dry seasons from four ecological settings in Cambodia (villages, forested areas near villages, rubber tree plantations and forest sites). Using odor-baited traps, 81% of the total samples across all sites were collected in cow baited traps, although 67% of the samples attracted by human baited traps were collected in forest sites. Overall, 20% of collected Anopheles were active during the day, with increased day biting during the dry season. 3131 samples were identified morphologically as 14 different species, and a subset was also identified by DNA amplicon sequencing allowing determination of 29 Anopheles species. The investigation of well characterized insecticide mutations (ace-1, kdr, and rdl genes) indicated that individuals carried mutations associated with response to all the different classes of insecticides. There also appeared to be a non-random association between mosquito species and insecticide resistance with Anopheles peditaeniatus exhibiting nearly fixed mutations. Molecular screening for Plasmodium sp. presence indicated that 3.6% of collected Anopheles were positive, most for P. vivax followed by P. falciparum. These results highlight some of the key mechanisms driving residual human malaria transmission in Cambodia, and illustrate the importance of diverse collection methods, sites and seasons to avoid bias and better characterize Anopheles mosquito ecology in Southeast Asia.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- World Health Organization. World malaria report 2020. 299 (2020).
-
- Leang R, et al. Evidence of Plasmodium falciparum malaria multidrug resistance to artemisinin and piperaquine in Western Cambodia: Dihydroartemisinin-piperaquine open-label multicenter clinical assessment. Antimicrob. Agents Chemother. 2015;59:4719–4726. doi: 10.1128/aac.00835-15. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
