

Chronic infection drives Dnmt3a-loss-of-function clonal hematopoiesis via IFNγ signaling
- PMID: 33743191
- PMCID: PMC8349829
- DOI: 10.1016/j.stem.2021.03.002
Chronic infection drives Dnmt3a-loss-of-function clonal hematopoiesis via IFNγ signaling
Abstract
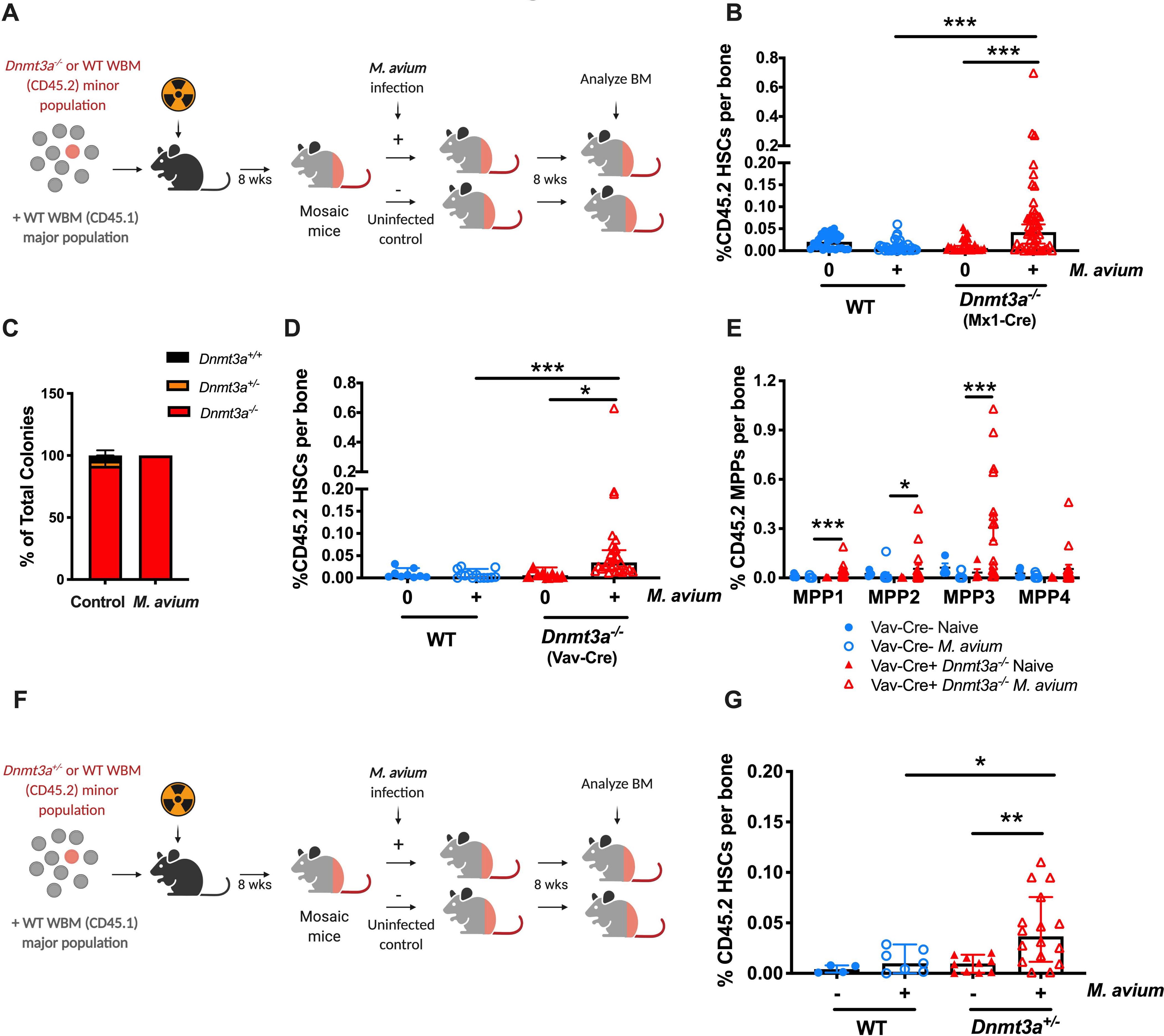
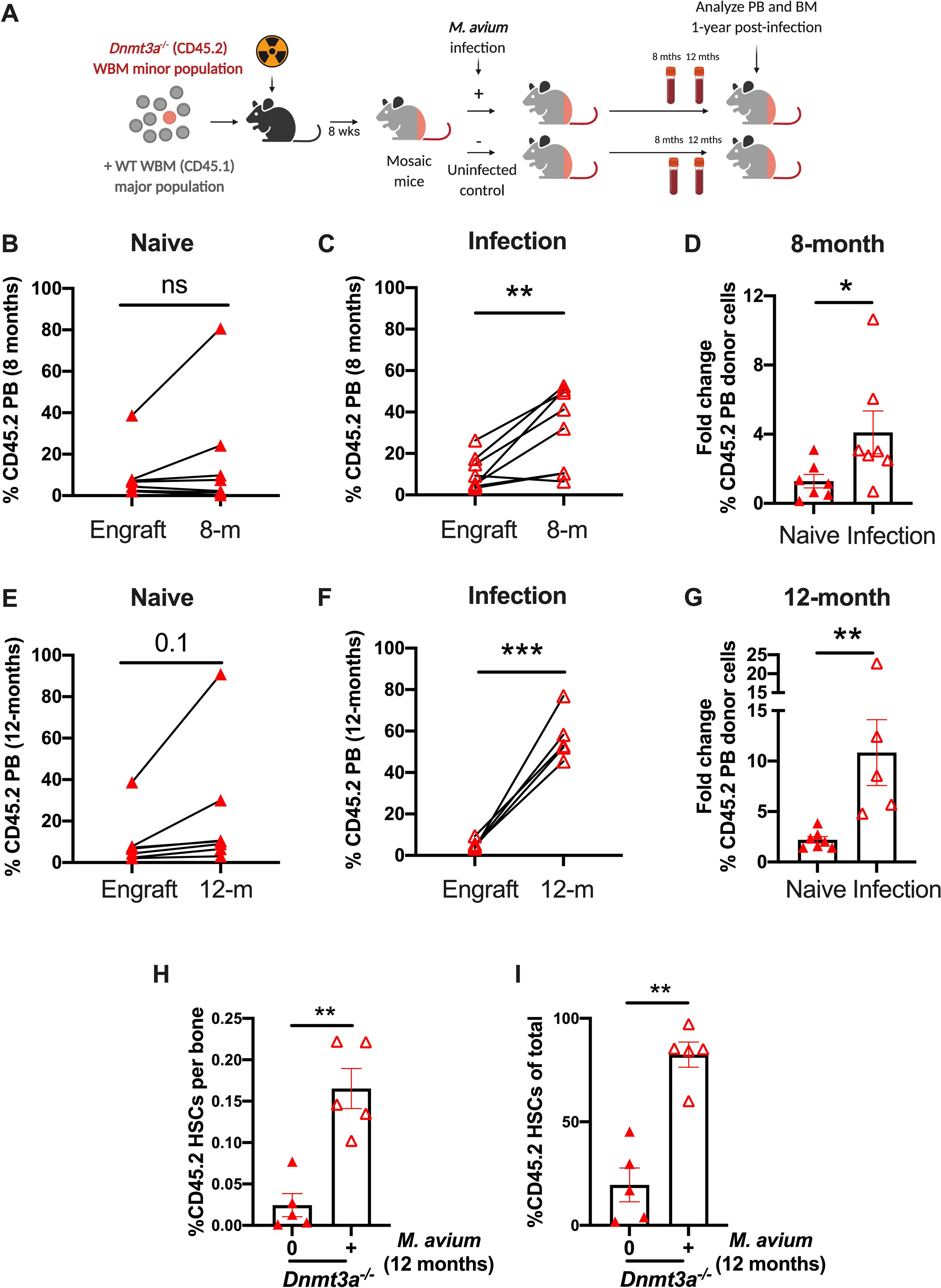
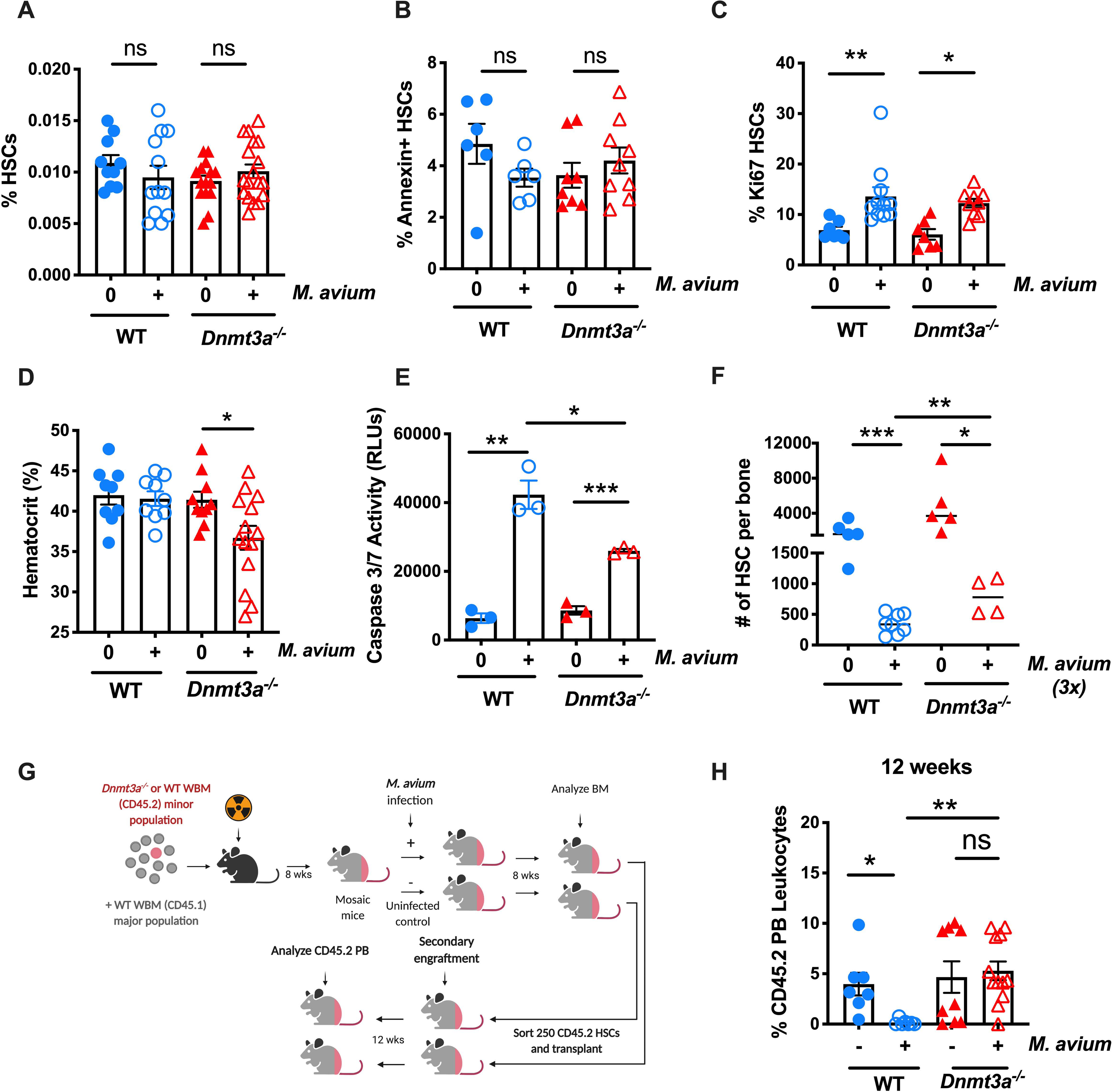
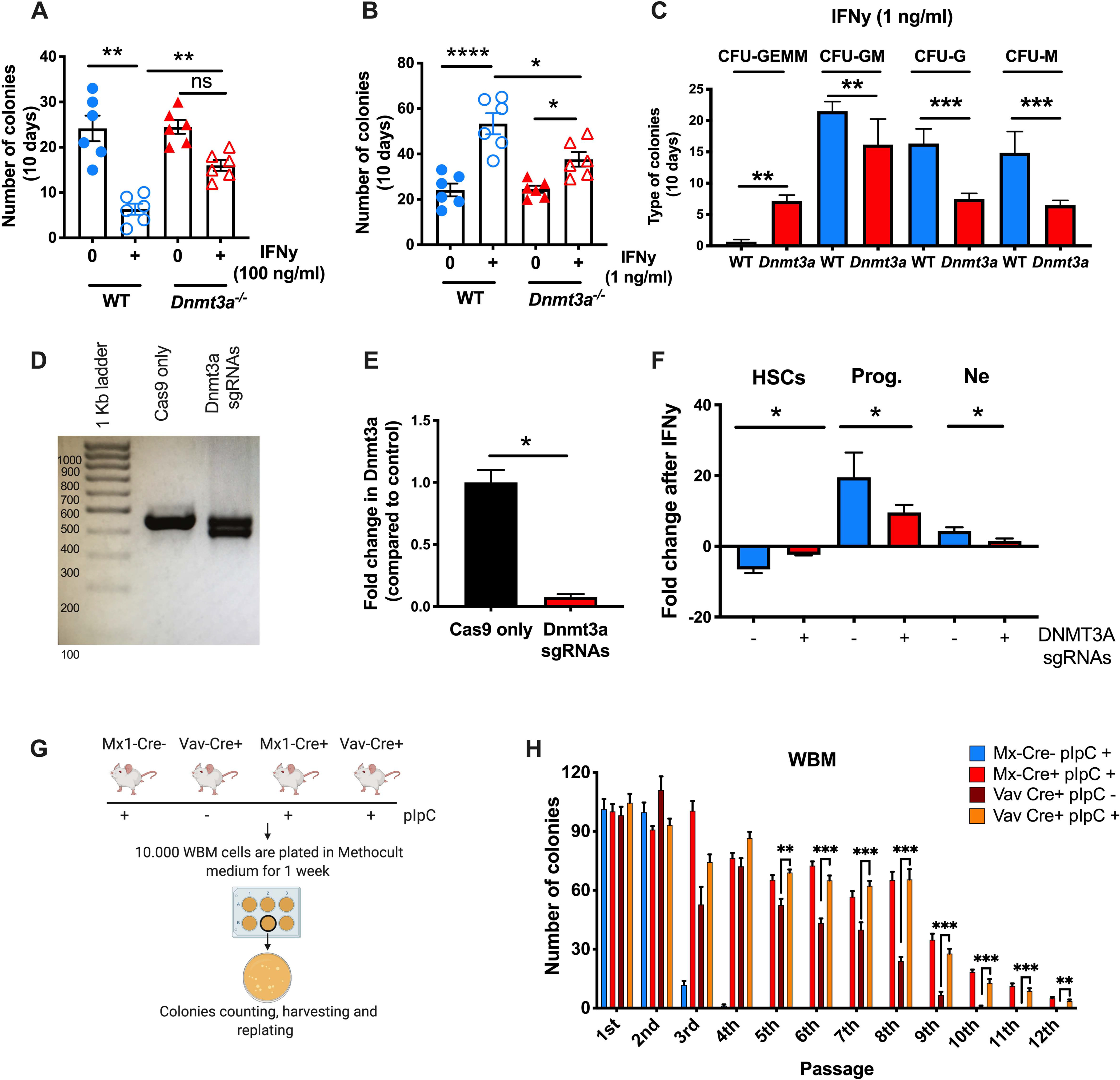
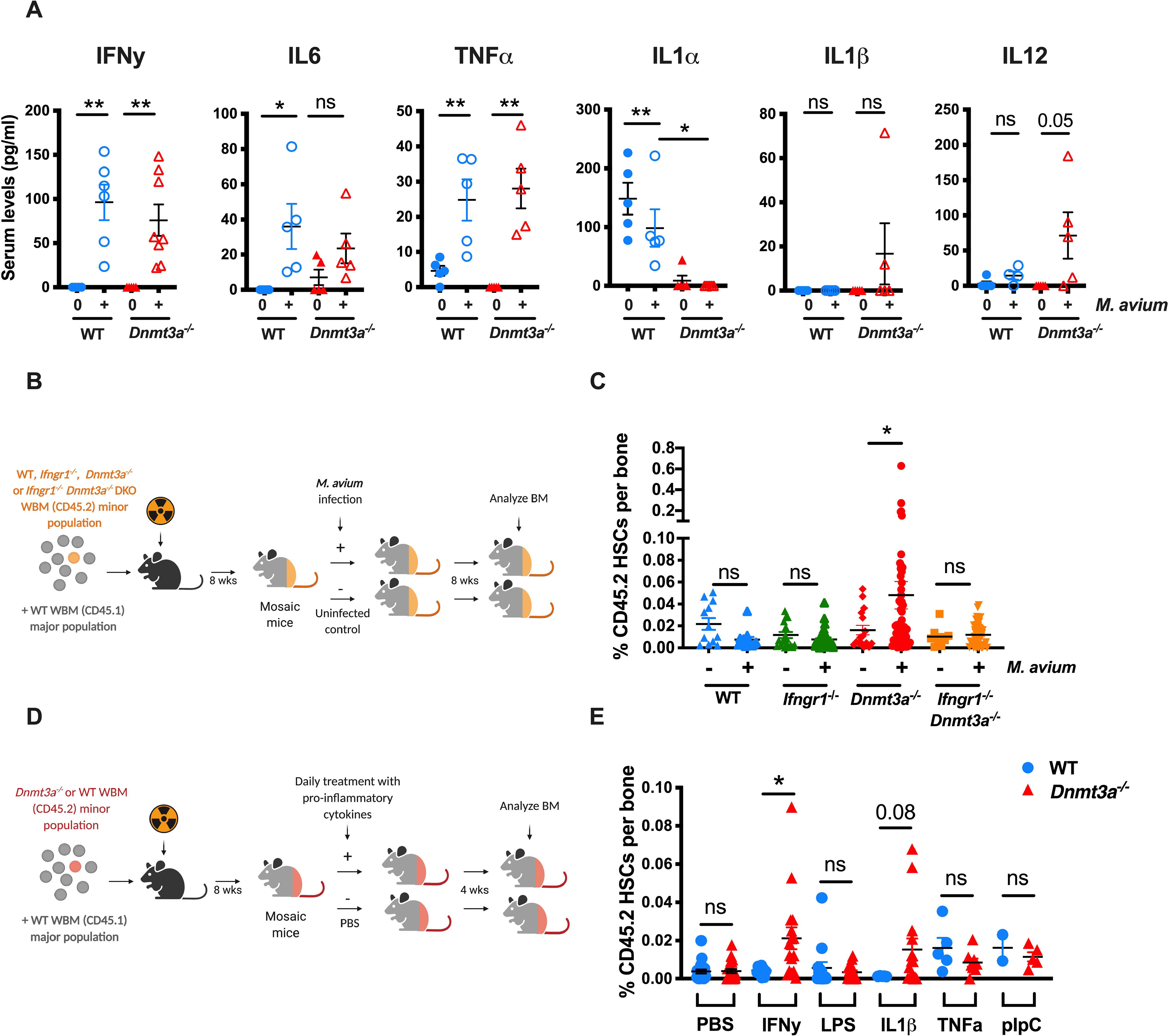
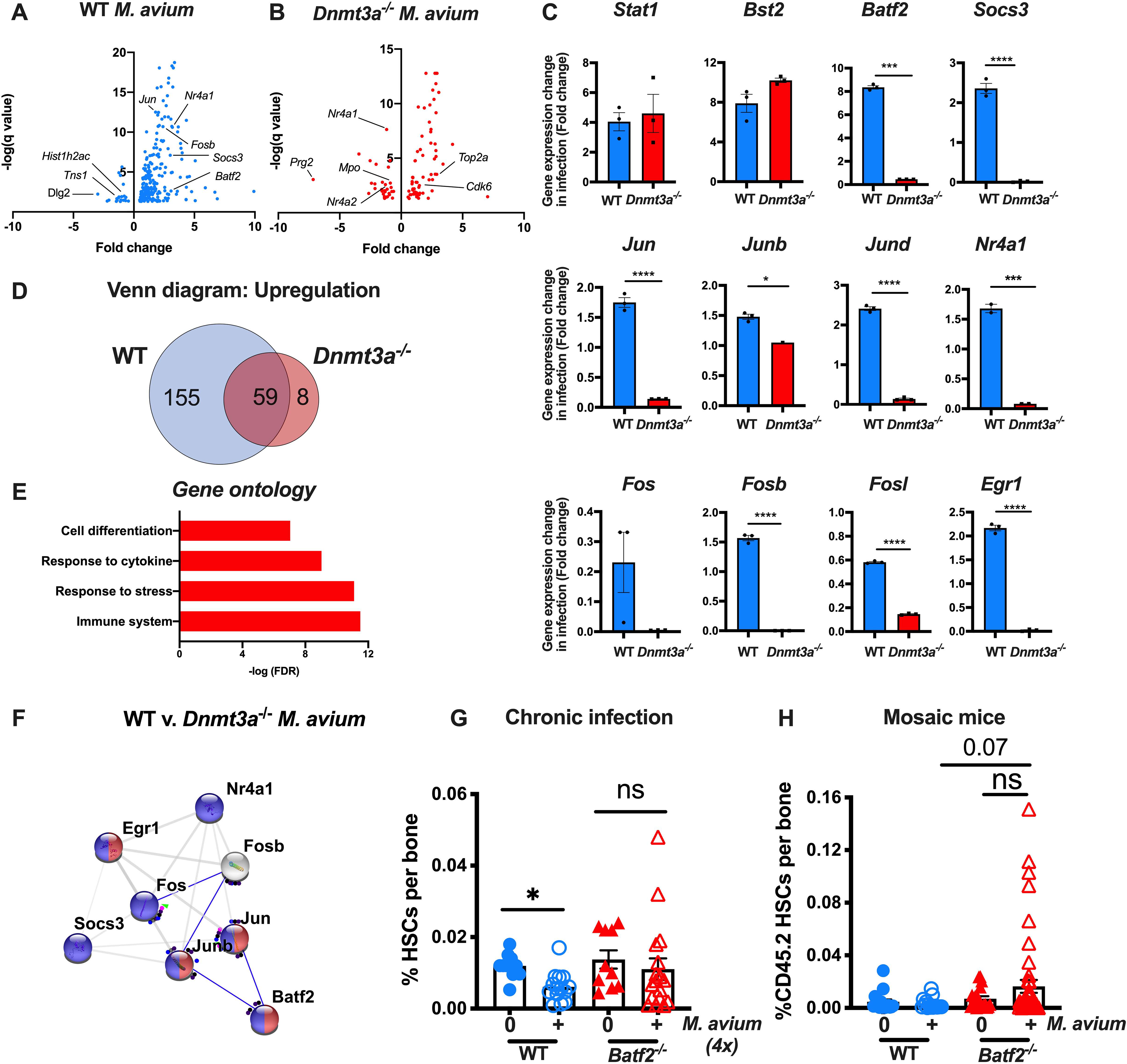
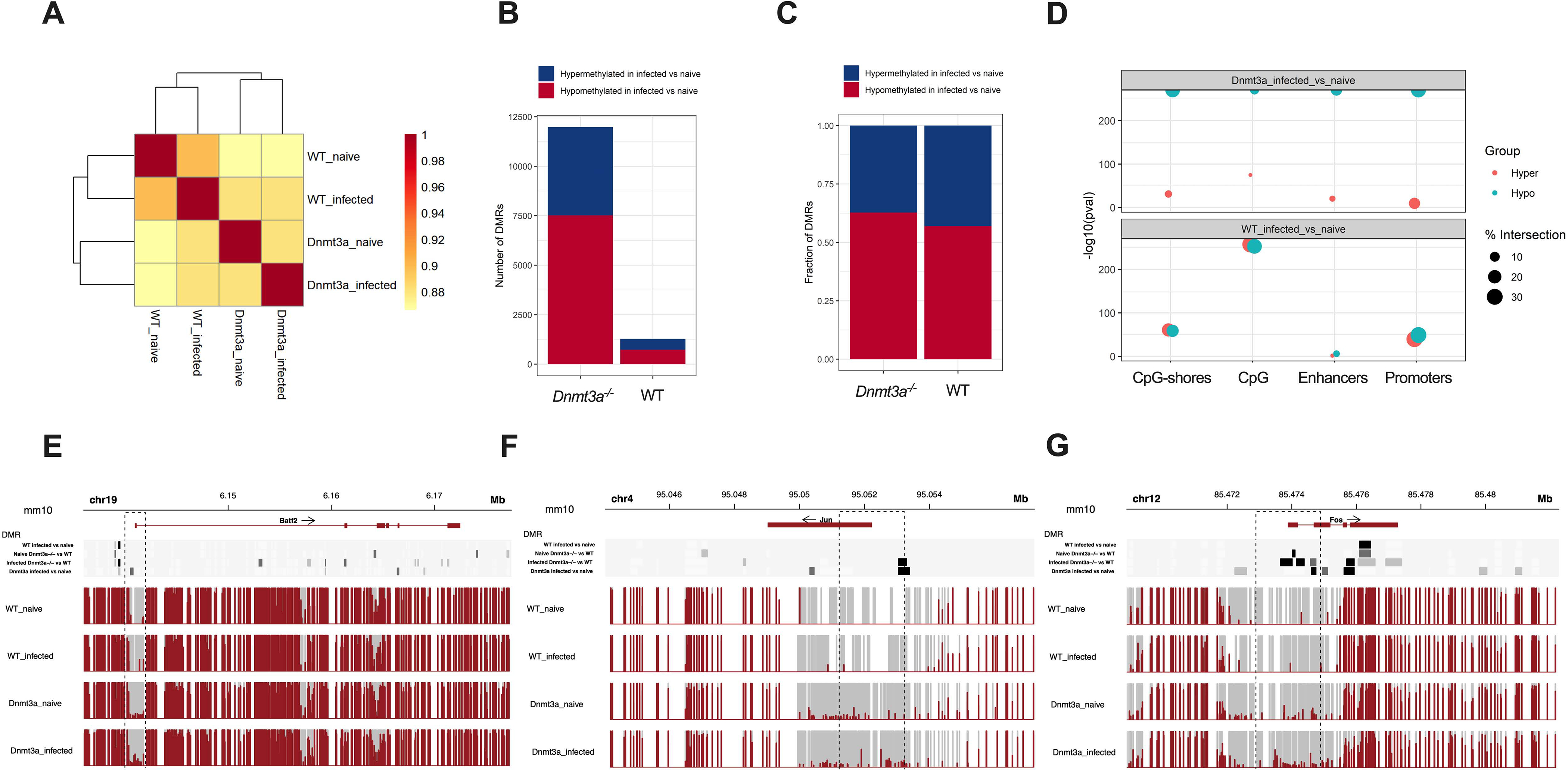
Age-related clonal hematopoiesis (CH) is a risk factor for malignancy, cardiovascular disease, and all-cause mortality. Somatic mutations in DNMT3A are drivers of CH, but decades may elapse between the acquisition of a mutation and CH, suggesting that environmental factors contribute to clonal expansion. We tested whether infection provides selective pressure favoring the expansion of Dnmt3a mutant hematopoietic stem cells (HSCs) in mouse chimeras. We created Dnmt3a-mosaic mice by transplanting Dnmt3a-/- and WT HSCs into WT mice and observed the substantial expansion of Dnmt3a-/- HSCs during chronic mycobacterial infection. Injection of recombinant IFNγ alone was sufficient to phenocopy CH by Dnmt3a-/- HSCs upon infection. Transcriptional and epigenetic profiling and functional studies indicate reduced differentiation associated with widespread methylation alterations, and reduced secondary stress-induced apoptosis accounts for Dnmt3a-/- clonal expansion during infection. DNMT3A mutant human HSCs similarly exhibit defective IFNγ-induced differentiation. We thus demonstrate that IFNγ signaling induced during chronic infection can drive DNMT3A-loss-of-function CH.
Keywords: CH; DNMT3A; bone marrow; clonal competition; clonal hematopoiesis; epigenetic regulation; hematopoietic stem cell; infection; interferon gamma; mycobacterial infection.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Infection makes micro-CHIPs into macro-CHIPs.Cell Stem Cell. 2021 Aug 5;28(8):1335-1336. doi: 10.1016/j.stem.2021.07.006. Cell Stem Cell. 2021. PMID: 34358435
References
-
- Abegunde SO, Buckstein R, Wells RA, and Rauh MJ (2018). An inflammatory environment containing TNFα favors Tet2-mutant clonal hematopoiesis. Experimental Hematology 59, 60–65. - PubMed
-
- Andrews S (2010). FastQC: a quality control tool for high throughput sequence data.
-
- Azrakhsh NA, Mensah-Glanowska P, Sand K, and Kittang AO (2019). Targeting Immune Signaling Pathways in Clonal Hematopoiesis. Current medicinal chemistry 26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F31 HL154661/HL/NHLBI NIH HHS/United States
- R35 HL155672/HL/NHLBI NIH HHS/United States
- S10 OD020066/OD/NIH HHS/United States
- P30 DK056338/DK/NIDDK NIH HHS/United States
- S10 RR024574/RR/NCRR NIH HHS/United States
- T32 DK060445/DK/NIDDK NIH HHS/United States
- R01 HL136333/HL/NHLBI NIH HHS/United States
- T32 HL092332/HL/NHLBI NIH HHS/United States
- P30 CA125123/CA/NCI NIH HHS/United States
- T32 AI053831/AI/NIAID NIH HHS/United States
- P30 AI036211/AI/NIAID NIH HHS/United States
- R01 HL134880/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
