

Beyond the double helix: DNA structural diversity and the PDB
- PMID: 33744292
- PMCID: PMC8063756
- DOI: 10.1016/j.jbc.2021.100553
Beyond the double helix: DNA structural diversity and the PDB
Abstract
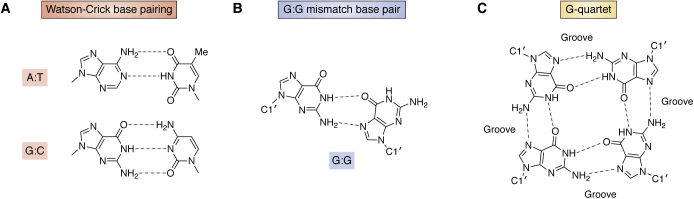
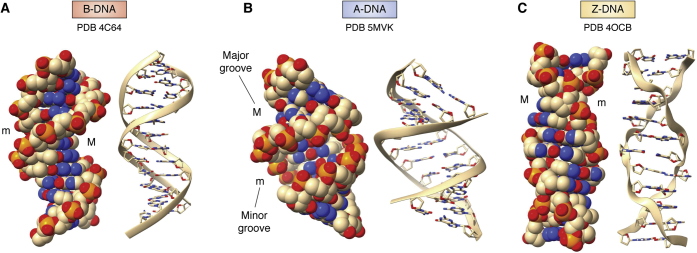
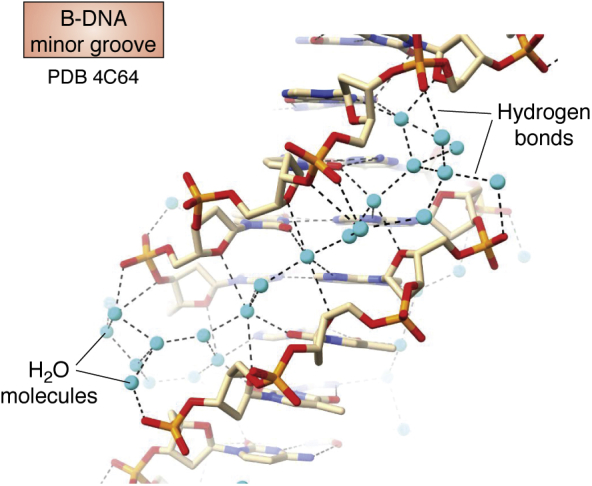
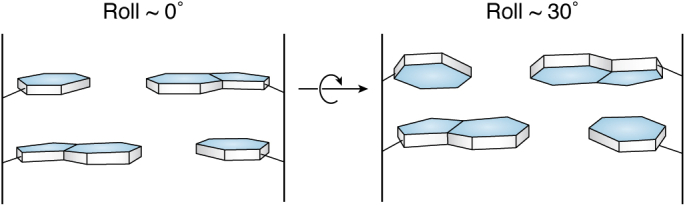
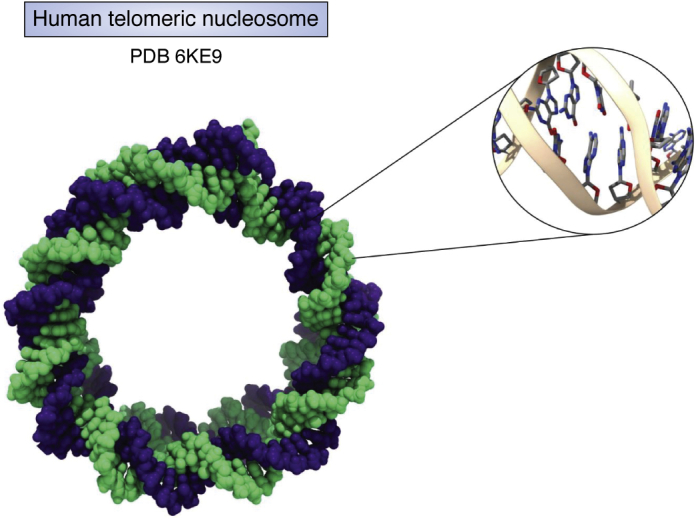
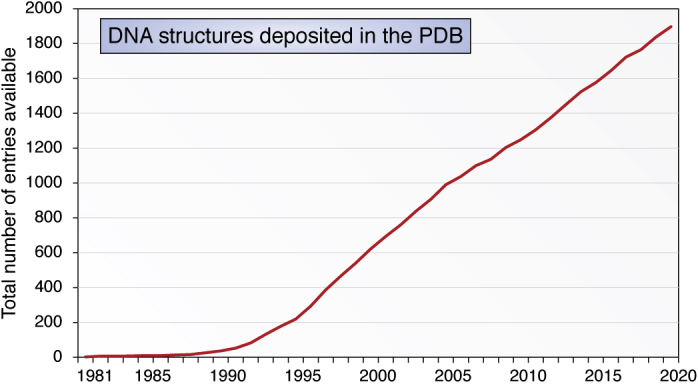
The determination of the double helical structure of DNA in 1953 remains the landmark event in the development of modern biological and biomedical science. This structure has also been the starting point for the determination of some 2000 DNA crystal structures in the subsequent 68 years. Their structural diversity has extended to the demonstration of sequence-dependent local structure in duplex DNA, to DNA bending in short and long sequences and in the DNA wound round the nucleosome, and to left-handed duplex DNAs. Beyond the double helix itself, in circumstances where DNA sequences are or can be induced to unwind from being duplex, a wide variety of topologies and forms can exist. Quadruplex structures, based on four-stranded cores of stacked G-quartets, are prevalent though not randomly distributed in the human and other genomes and can play roles in transcription, translation, and replication. Yet more complex folds can result in DNAs with extended tertiary structures and enzymatic/catalytic activity. The Protein Data Bank is the depository of all these structures, and the resource where structures can be critically examined and validated, as well as compared one with another to facilitate analysis of conformational and base morphology features. This review will briefly survey the major structural classes of DNAs and illustrate their significance, together with some examples of how the use of the Protein Data Bank by for example, data mining, has illuminated DNA structural concepts.
Keywords: DNA; NDB; NMR; PDB; crystal structures; deoxyoligonucleotides; double helix; multistrand helices; quadruplex; sequence-dependent structure.
Copyright © 2021 The Author. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The author declares that he has no conflicts of interest with the contents of this manuscript.
Figures
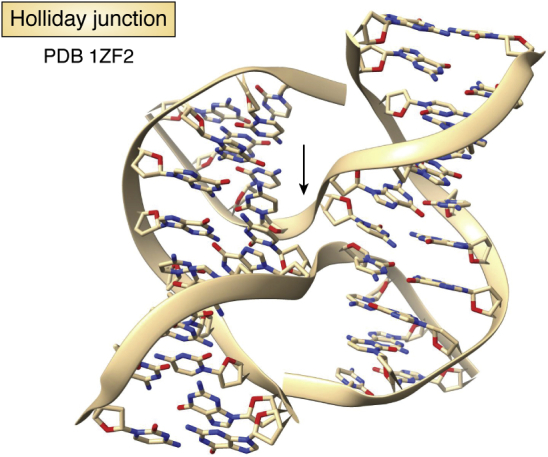
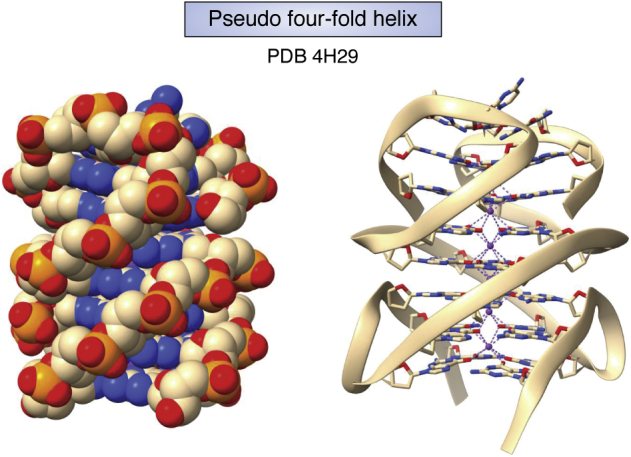
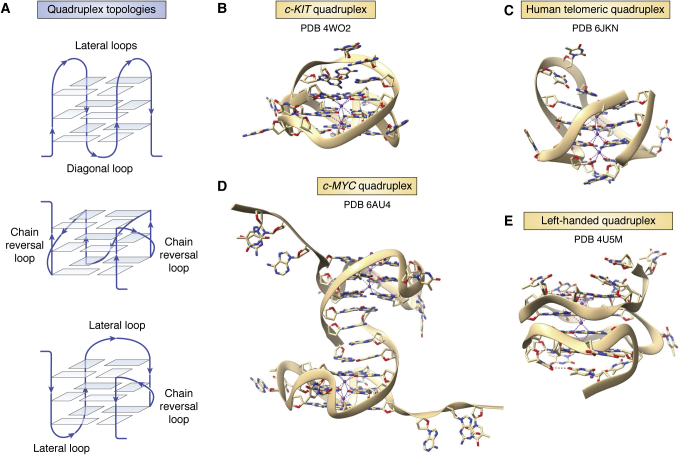
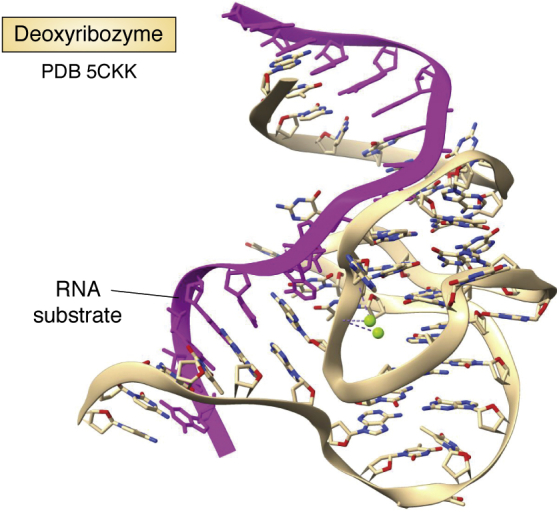
References
-
- Neidle S. Academic Press; London: 2008. Principles of Nucleic Acid Structure.
-
- Watson J.D., Crick F.H. Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature. 1953;171:737–738. - PubMed
-
- Wilkins M.H., Stokes A.R., Wilson H.R. Molecular structure of deoxypentose nucleic acids. Nature. 1953;171:738–740. - PubMed
-
- Franklin R.E., Gosling R.G. Molecular configuration in sodium thymonucleate. Nature. 1953;171:740–741. - PubMed
-
- Arnott S., Hutchinson F., Spencer M., Wilkins M.H., Fuller W., Langridge R. X-ray diffraction studies of double helical ribonucleic acid. Nature. 1966;211:227–232. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
