

Retroviral prototype foamy virus intasome binding to a nucleosome target does not determine integration efficiency
- PMID: 33744295
- PMCID: PMC8050864
- DOI: 10.1016/j.jbc.2021.100550
Retroviral prototype foamy virus intasome binding to a nucleosome target does not determine integration efficiency
Abstract
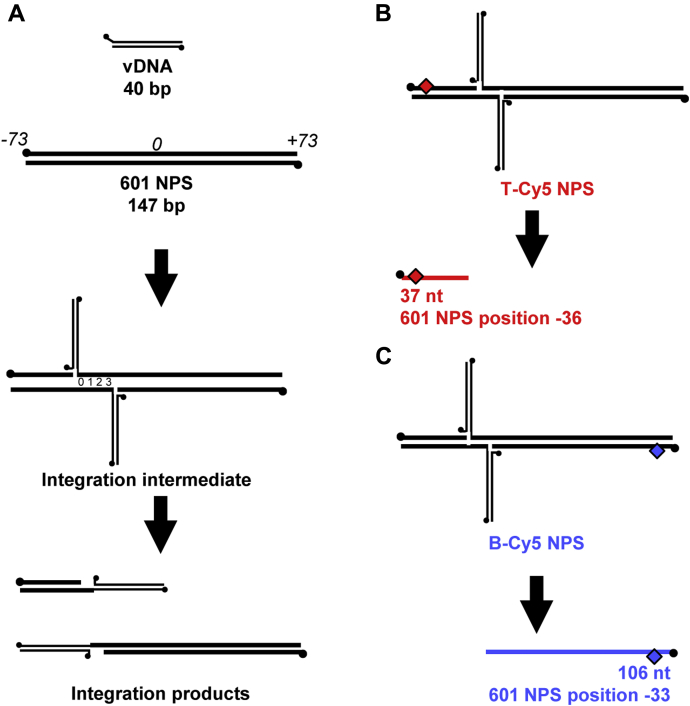
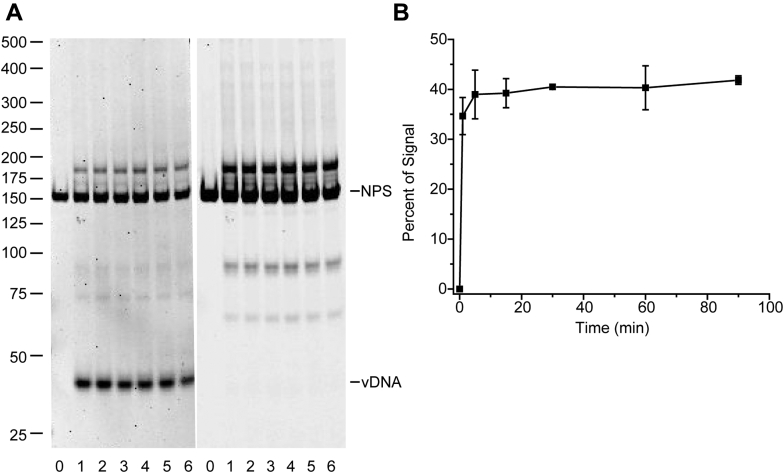
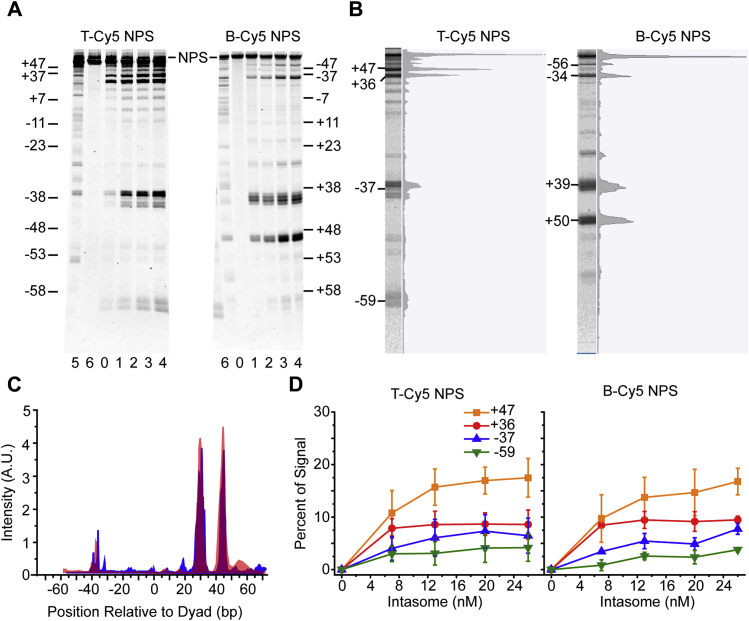
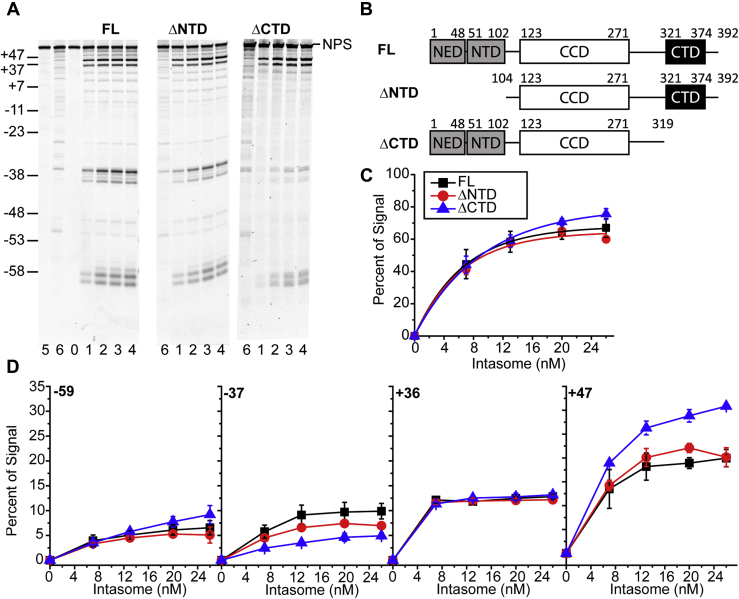
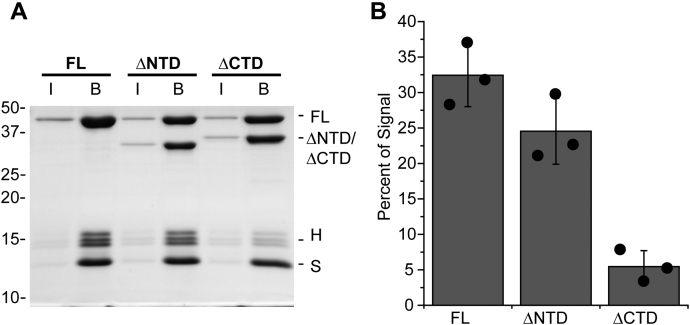
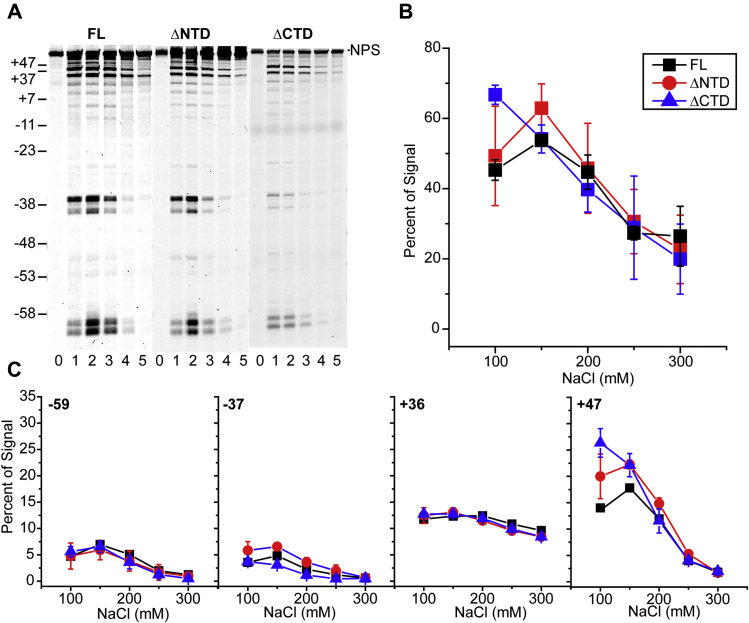
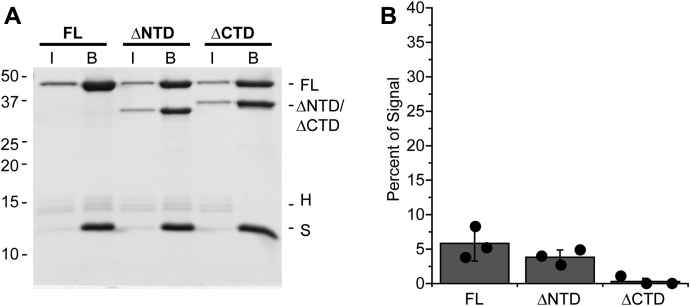
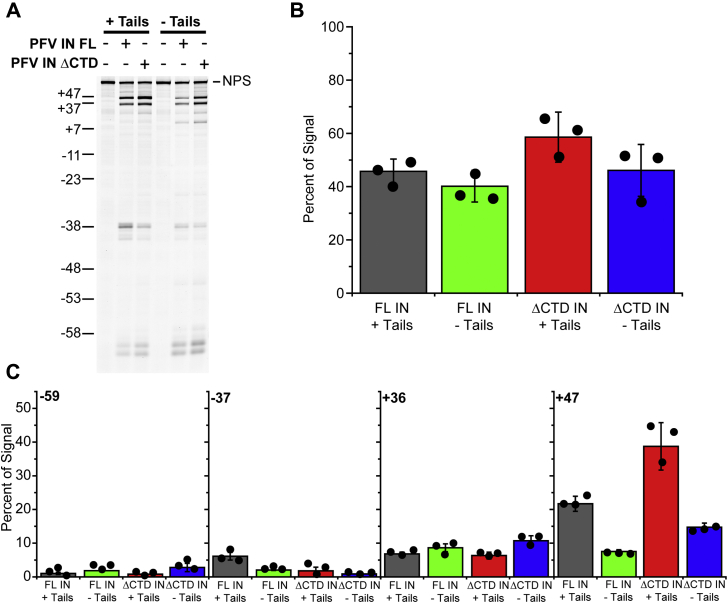
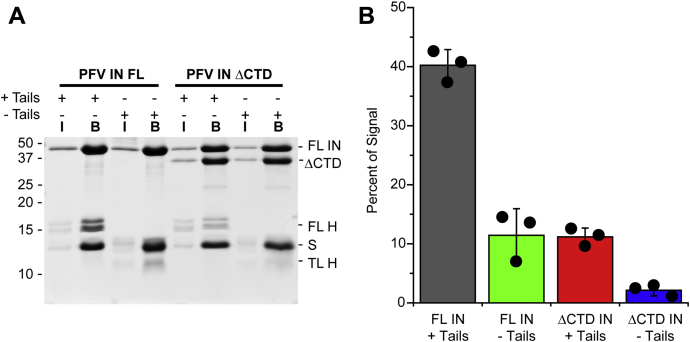
Retroviral integrases must navigate host DNA packaged as chromatin during integration of the viral genome. Prototype foamy virus (PFV) integrase (IN) forms a tetramer bound to two viral DNA (vDNA) ends in a complex termed an intasome. PFV IN consists of four domains: the amino terminal extension domain (NED), amino terminal domain (NTD), catalytic core domain (CCD), and carboxyl terminal domain (CTD). The domains of the two inner IN protomers have been visualized, as well as the CCDs of the two outer IN protomers. However, the roles of the amino and carboxyl terminal domains of the PFV intasome outer subunits during integration to a nucleosome target substrate are not clear. We used the well-characterized 601 nucleosome to assay integration activity as well as intasome binding. PFV intasome integration to 601 nucleosomes occurs in clusters at four independent sites. We find that the outer protomer NED and NTD domains have no significant effects on integration efficiency, site selection, or binding. The CTDs of the outer PFV intasome subunits dramatically affect nucleosome binding but have little effect on total integration efficiency. The outer PFV IN CTDs did significantly alter the integration efficiency at one site. Histone tails also significantly affect intasome binding, but have little impact on PFV integration efficiency or site selection. These results indicate that binding to nucleosomes does not correlate with integration efficiency and suggests most intasome-binding events are unproductive.
Keywords: enzyme catalysis; integration; nucleosome; protein–protein interaction; prototype foamy virus; retrovirus.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Coffin J.M., Hughes S.H., Varmus H.E. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 1997. Retroviruses. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
