

Complement Activation in the Central Nervous System: A Biophysical Model for Immune Dysregulation in the Disease State
- PMID: 33746710
- PMCID: PMC7969890
- DOI: 10.3389/fnmol.2021.620090
Complement Activation in the Central Nervous System: A Biophysical Model for Immune Dysregulation in the Disease State
Abstract
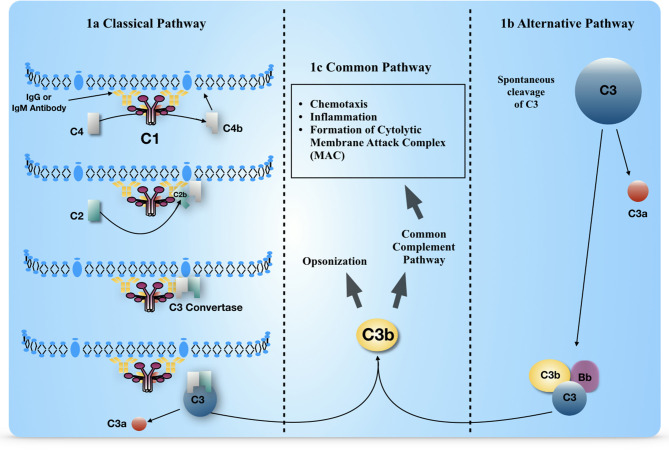
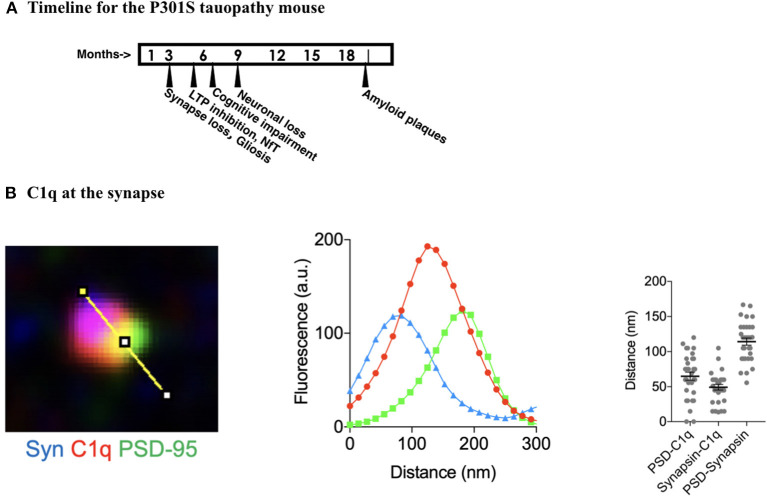
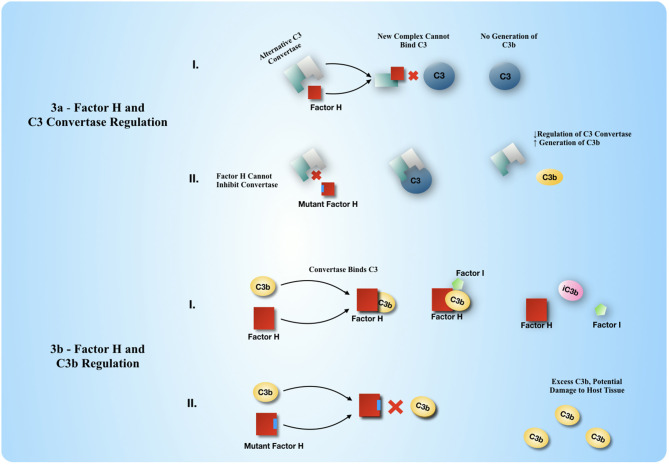
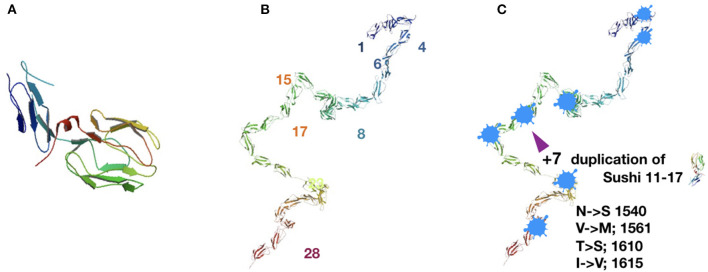
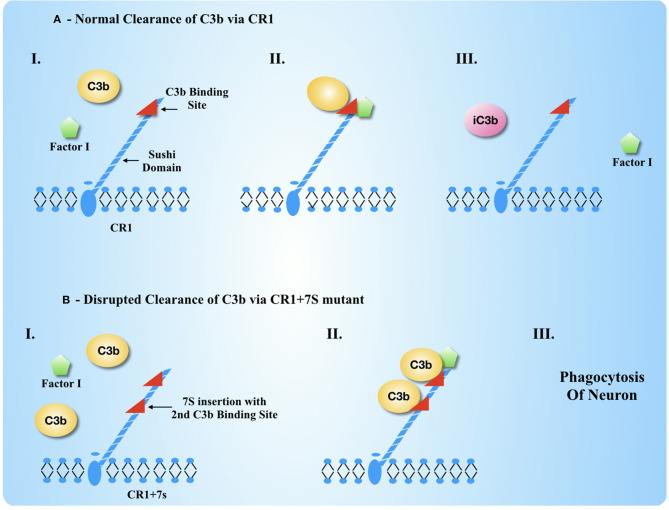
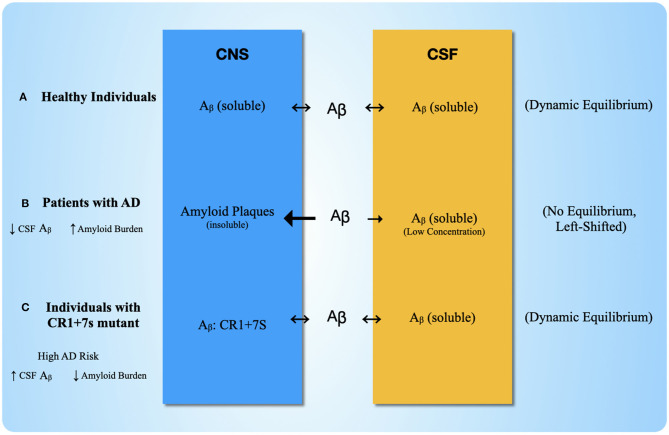
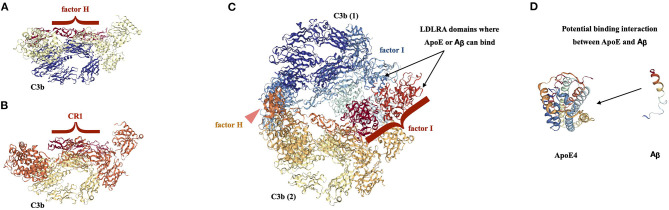
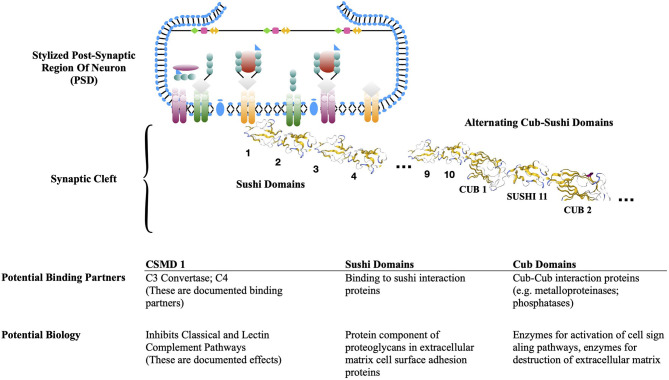
Complement, a feature of the innate immune system that targets pathogens for phagocytic clearance and promotes inflammation, is tightly regulated to prevent damage to host tissue. This regulation is paramount in the central nervous system (CNS) since complement proteins degrade neuronal synapses during development, homeostasis, and neurodegeneration. We propose that dysregulated complement, particularly C1 or C3b, may errantly target synapses for immune-mediated clearance, therefore highlighting regulatory failure as a major potential mediator of neurological disease. First, we explore the mechanics of molecular neuroimmune relationships for the regulatory proteins: Complement Receptor 1, C1-Inhibitor, Factor H, and the CUB-sushi multiple domain family. We propose that biophysical and chemical principles offer clues for understanding mechanisms of dysregulation. Second, we describe anticipated effects to CNS disease processes (particularly Alzheimer's Disease) and nest our ideas within existing basic science, clinical, and epidemiological findings. Finally, we illustrate how the concepts presented within this manuscript provoke new ways of approaching age-old neurodegenerative processes. Every component of this model is testable by straightforward experimentation and highlights the untapped potential of complement dysregulation as a driver of CNS disease. This includes a putative role for complement-based neurotherapeutic agents and companion biomarkers.
Keywords: Alzheimer's disease; C1 inhibitor; CR1; complement; factor H; neuroimmune; schizophrenia.
Copyright © 2021 Peoples and Strang.
Conflict of interest statement
CS is an unpaid consultant to IPPIN Biomarkers and will become a shareholder of the C-Corporation should it obtain Series A financial backing. The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Abbas A. K., Lichtman A. H., Pillai S. (2019). Basic Immunology: Functions and Disorders of the Immune System. Philidelphia, PA: Elsevier Health Sciences.
LinkOut - more resources
Full Text Sources
Other Literature Sources