

Retracing Storage Polysaccharide Evolution in Stramenopila
- PMID: 33747010
- PMCID: PMC7965971
- DOI: 10.3389/fpls.2021.629045
Retracing Storage Polysaccharide Evolution in Stramenopila
Abstract
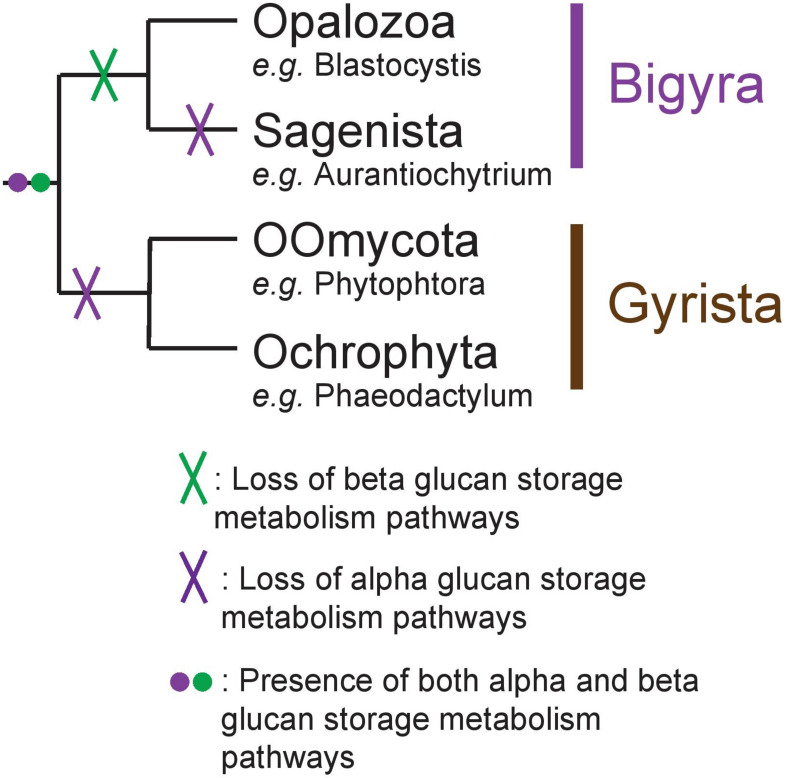
Eukaryotes most often synthesize storage polysaccharides in the cytosol or vacuoles in the form of either alpha (glycogen/starch)- or beta-glucosidic (chrysolaminarins and paramylon) linked glucan polymers. In both cases, the glucose can be packed either in water-soluble (glycogen and chrysolaminarins) or solid crystalline (starch and paramylon) forms with different impacts, respectively, on the osmotic pressure, the glucose accessibility, and the amounts stored. Glycogen or starch accumulation appears universal in all free-living unikonts (metazoa, fungi, amoebozoa, etc.), as well as Archaeplastida and alveolata, while other lineages offer a more complex picture featuring both alpha- and beta-glucan accumulators. We now infer the distribution of these polymers in stramenopiles through the bioinformatic detection of their suspected metabolic pathways. Detailed phylogenetic analysis of key enzymes of these pathways correlated to the phylogeny of Stramenopila enables us to retrace the evolution of storage polysaccharide metabolism in this diverse group of organisms. The possible ancestral nature of glycogen metabolism in eukaryotes and the underlying source of its replacement by beta-glucans are discussed.
Keywords: CAZy; glycogen; laminarin; metabolism; polysaccharide; stramenopila.
Copyright © 2021 Chabi, Leleu, Fermont, Colpaert, Colleoni, Ball and Cenci.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
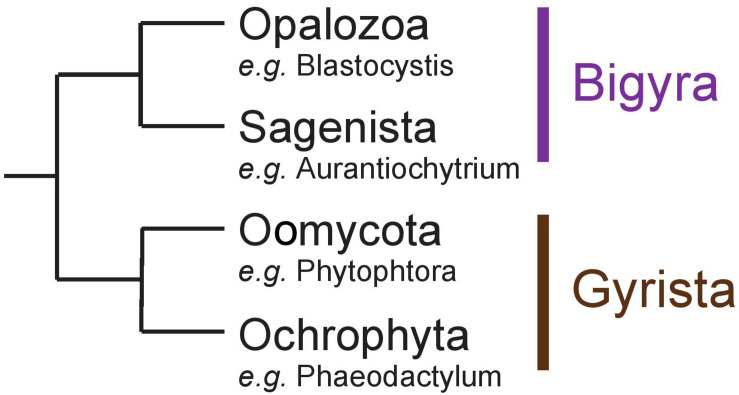
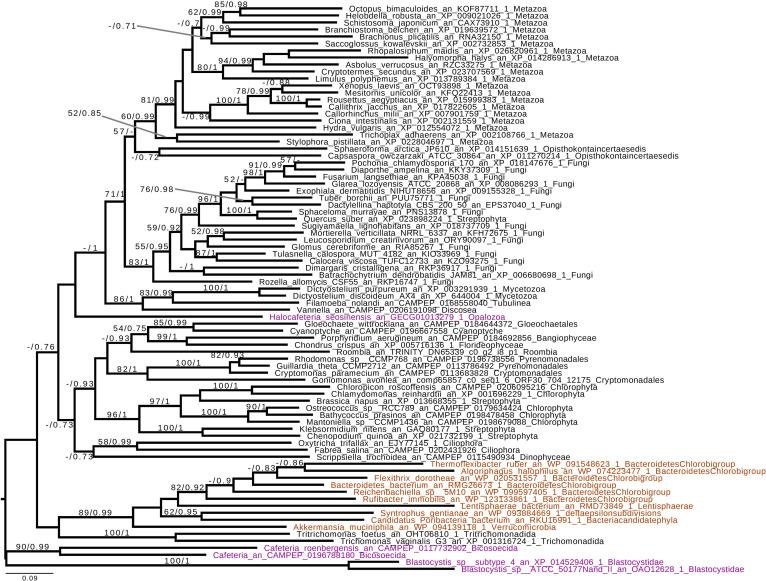
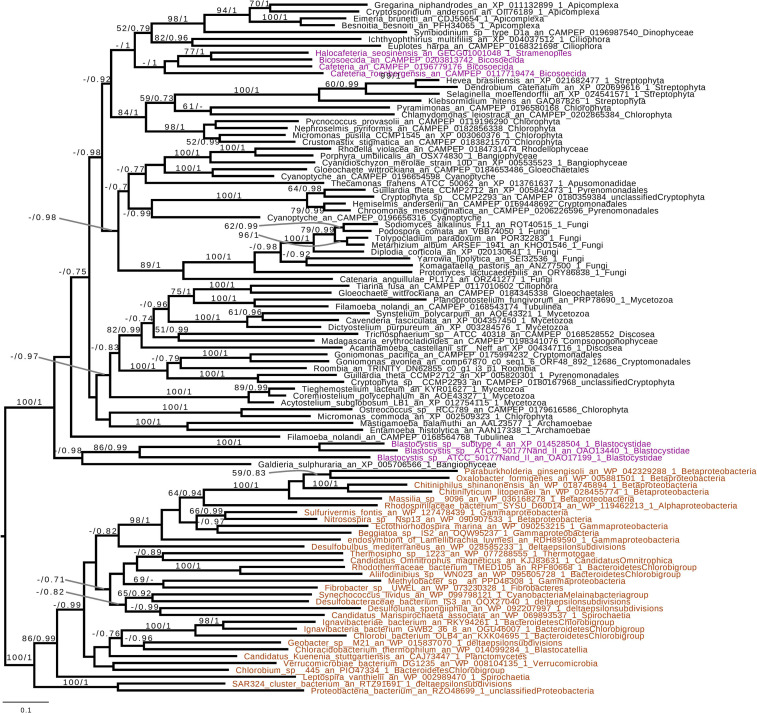
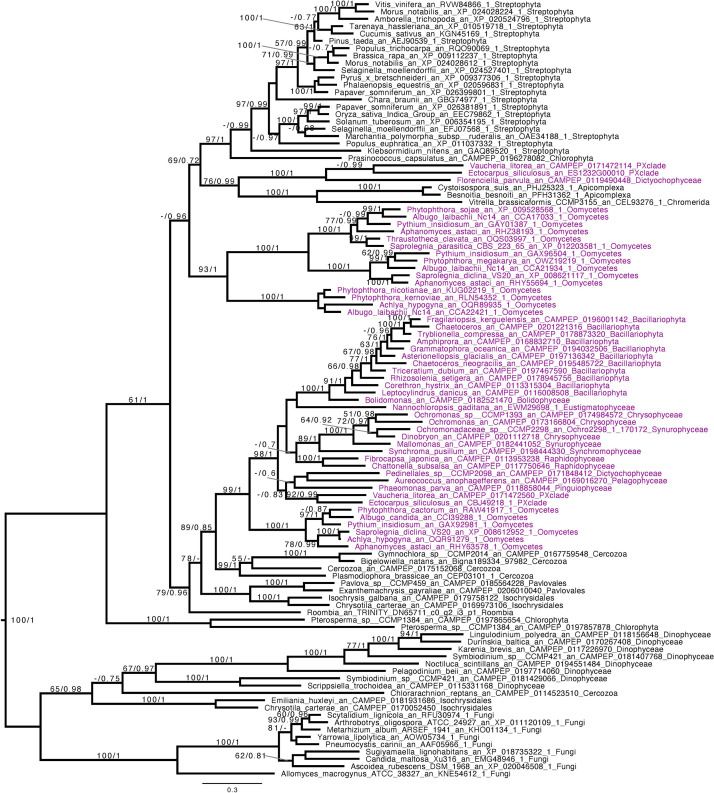

References
-
- Burki F., Kaplan M., Tikhonenkov D. V., Zlatogursky V., Minh B. Q., Radaykina L. V., et al. (2016). Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista. Proc. R. Soc. B Biol. Sci. 283 20152802. 10.1098/rspb.2015.2802 - DOI - PMC - PubMed
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources