

Evolution of Reproductive Division of Labor - Lessons Learned From the Social Amoeba Dictyostelium discoideum During Its Multicellular Development
- PMID: 33748102
- PMCID: PMC7969725
- DOI: 10.3389/fcell.2021.599525
Evolution of Reproductive Division of Labor - Lessons Learned From the Social Amoeba Dictyostelium discoideum During Its Multicellular Development
Abstract
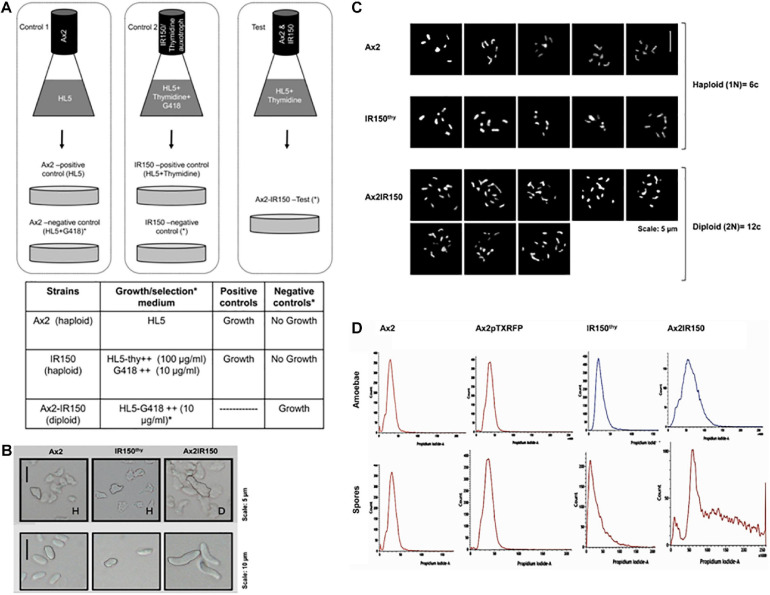
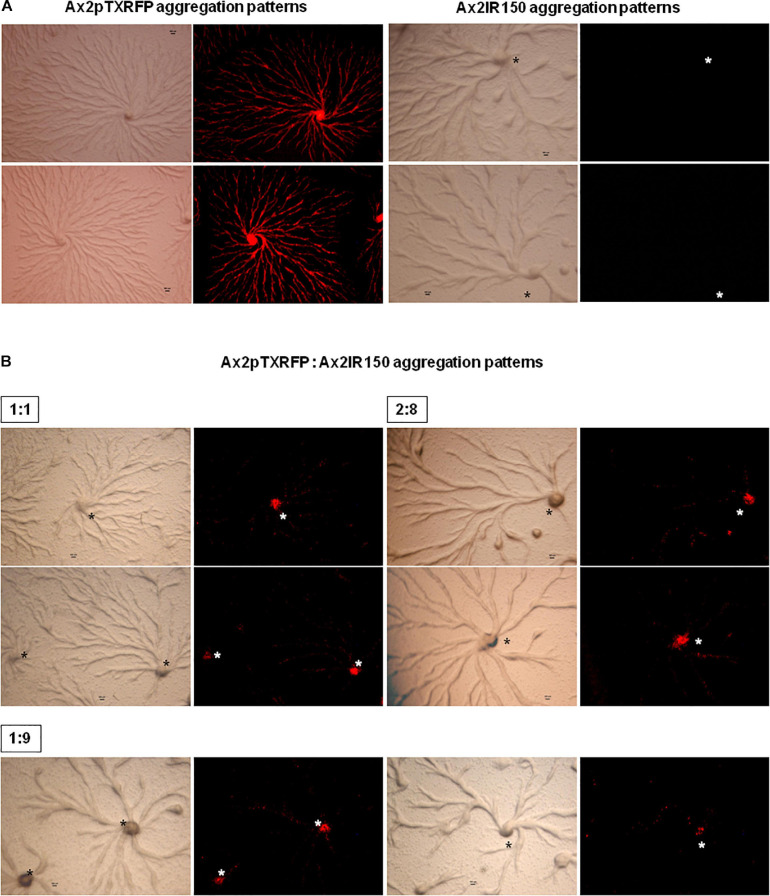
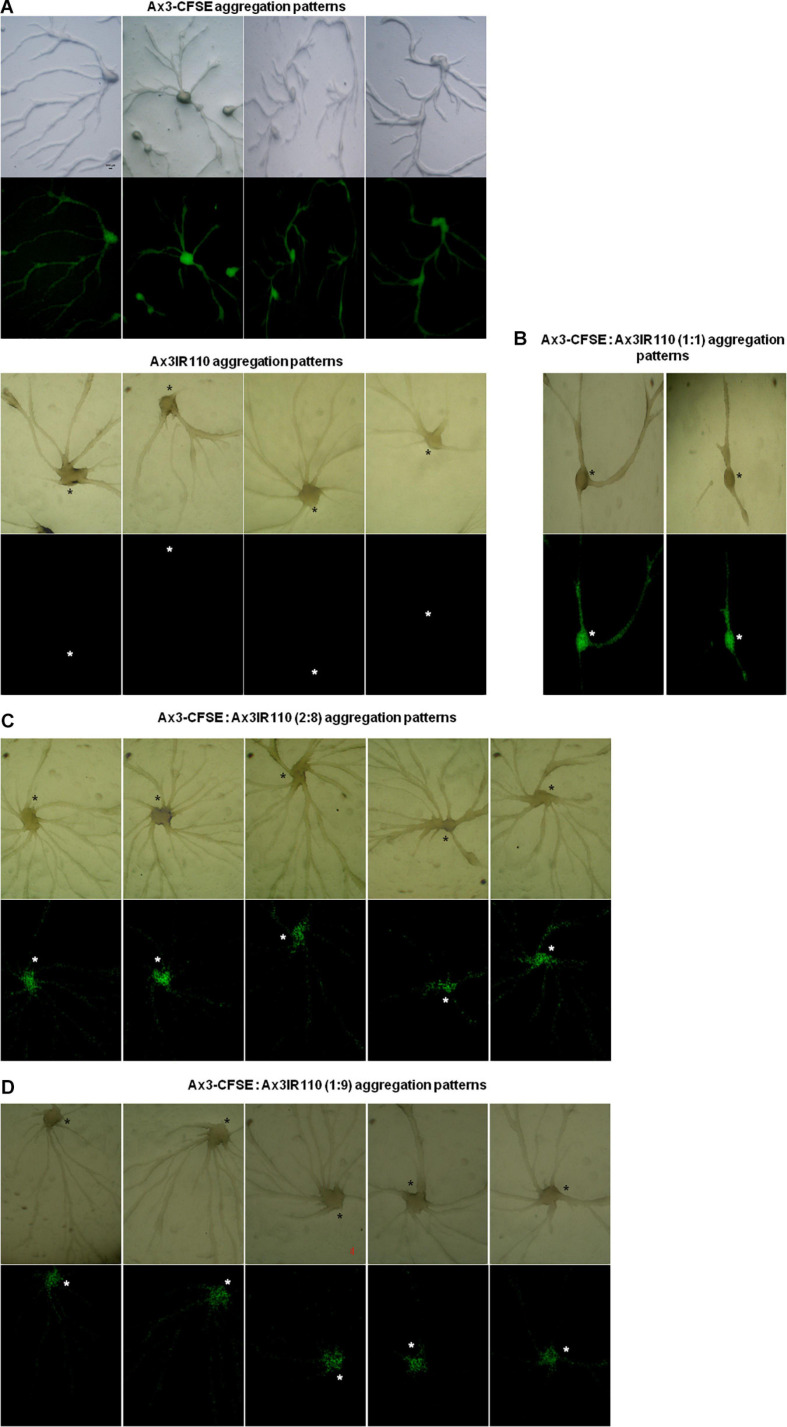
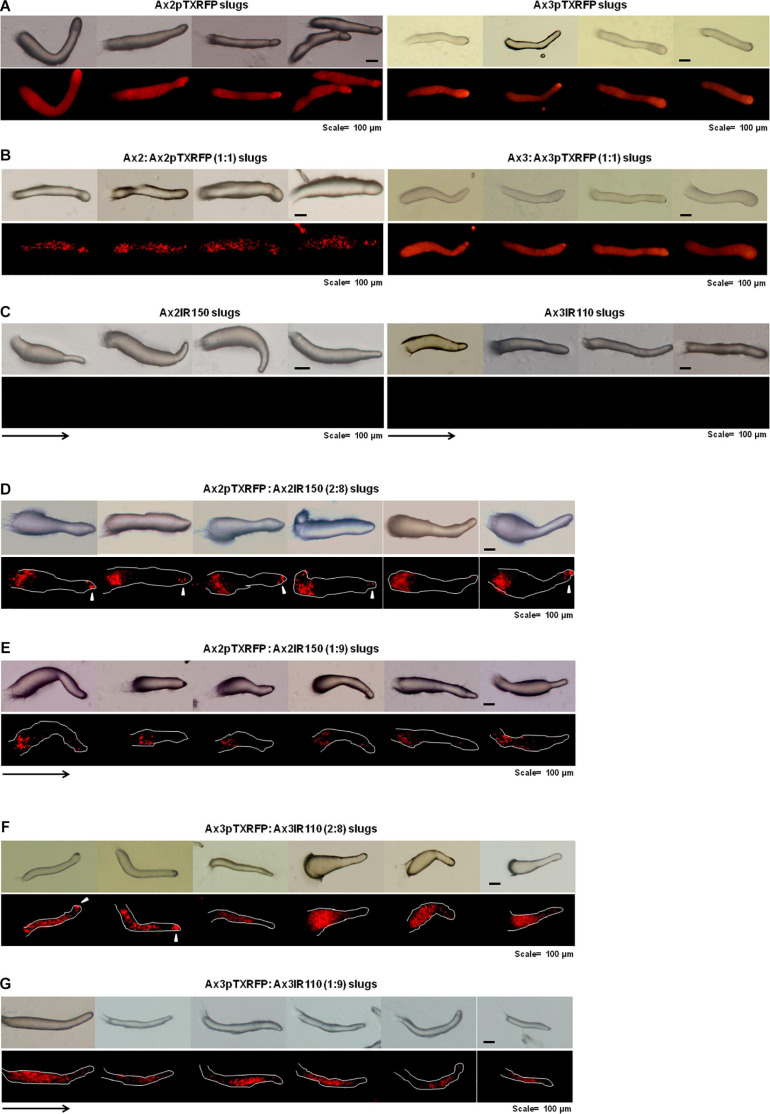
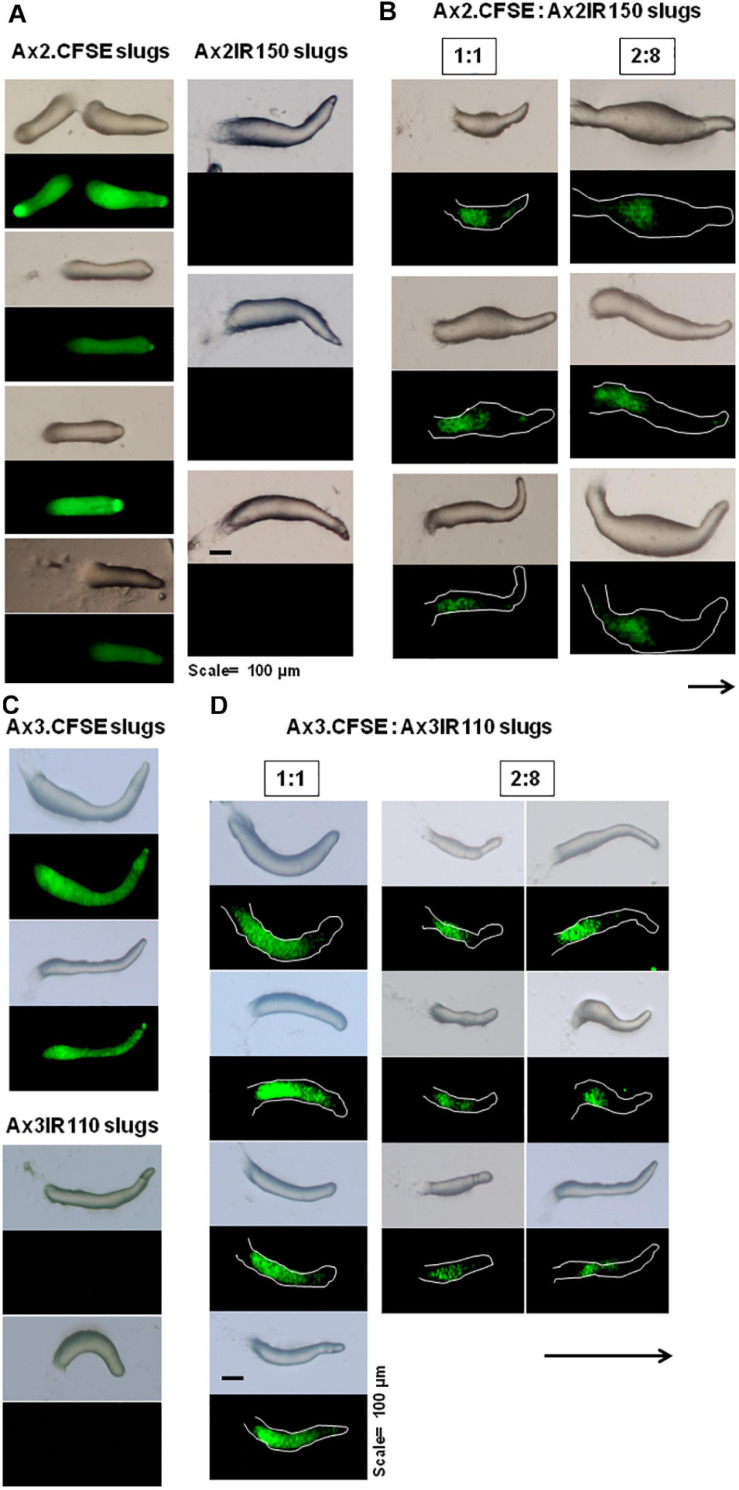
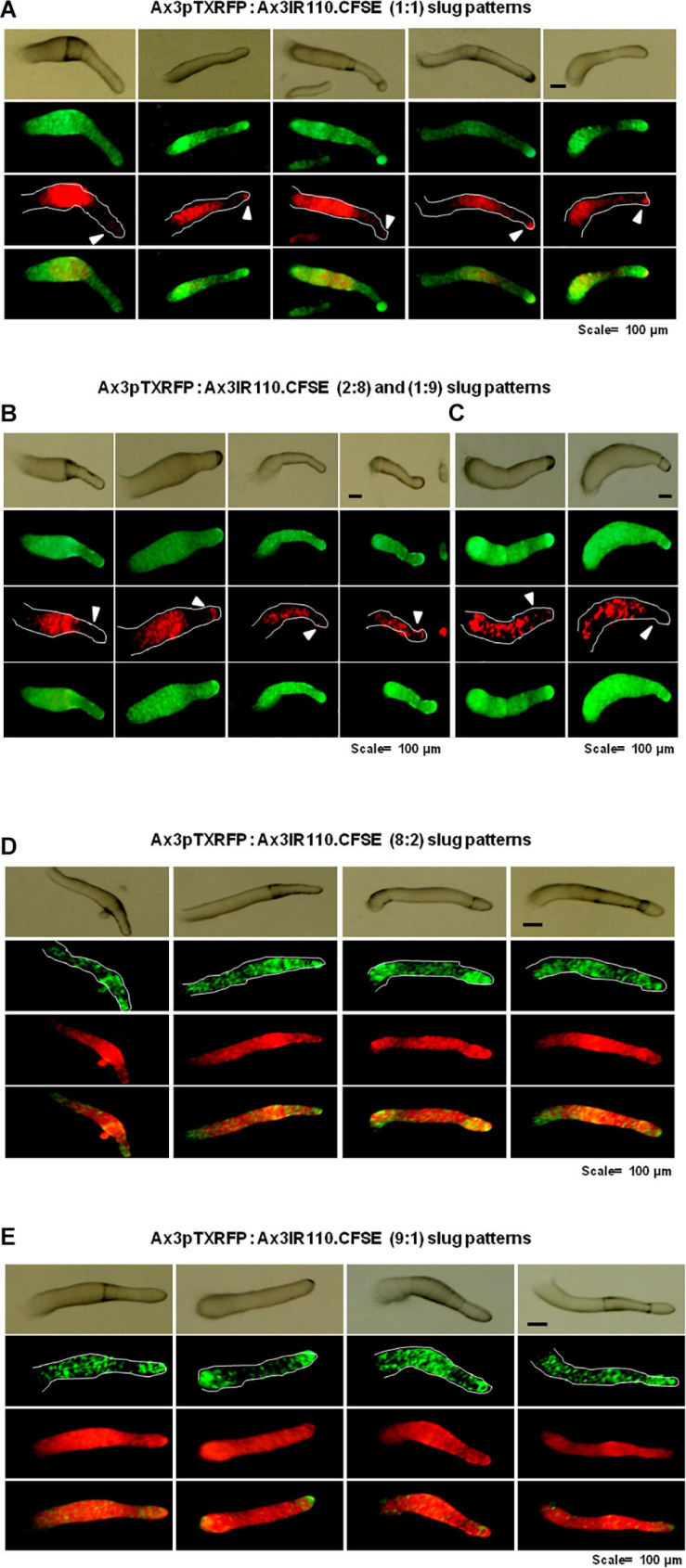
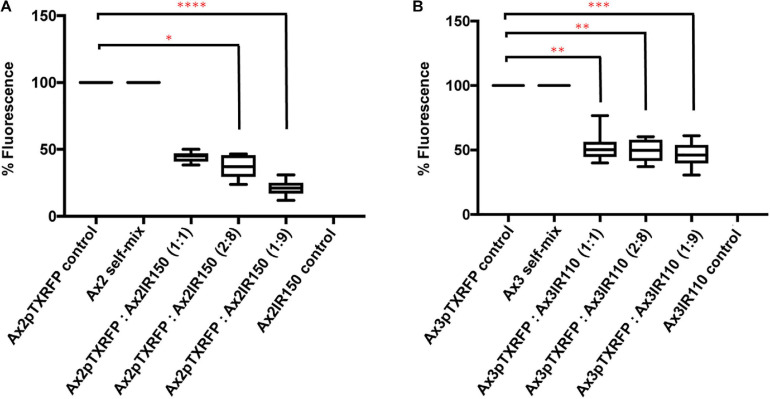
The origin of multicellular life from unicellular beings is an epochal step in the evolution of eukaryotes. There are several factors influencing cell fate choices during differentiation and morphogenesis of an organism. Genetic make-up of two cells that unite and fertilize is the key factor to signal the formation of various cell-types in due course of development. Although ploidy of the cell-types determines the genetics of an individual, the role of ploidy in cell fate decisions remains unclear. Dictyostelium serves as a versatile model to study the emergence of multicellular life from unicellular life forms. In this work, we investigate the role played by ploidy status of a cell on cell fate commitments during Dictyostelium development. To answer this question, we created Dictyostelium cells of different ploidy: haploid parents and derived isogenic diploids, allowing them to undergo development. The diploid strains used in this study were generated using parasexual genetics. The ploidy status of the haploids and diploids were confirmed by microscopy, flow cytometry, and karyotyping. Prior to reconstitution, we labeled the cells by two methods. First, intragenic expression of red fluorescent protein (RFP) and second, staining the amoebae with a vital, fluorescent dye carboxyfluorescein succinimidyl ester (CFSE). RFP labeled haploid cells allowed us to track the haploids in the chimeric aggregates, slugs, and fruiting bodies. The CFSE labeling method allowed us to track both the haploids and the diploids in the chimeric developmental structures. Our findings illustrate that the haploids demonstrate sturdy cell fate commitment starting from the aggregation stage. The haploids remain crowded at the aggregation centers of the haploid-diploid chimeric aggregates. At the slug stage haploids are predominantly occupying the slug posterior, and are visible in the spore population in the fruiting bodies. Our findings show that cell fate decisions during D. discoideum development are highly influenced by the ploidy status of a cell, adding a new aspect to already known factors Here, we report that ploidy status of a cell could also play a crucial role in regulating the cell fate commitments.
Keywords: Dictyostelium discoideum; aggregation multicellularity; cell fate choices; division multicellularity; ploidy; reproductive division of labor.
Copyright © 2021 Dhakshinamoorthy and Singh.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Investigation of cell patterning in the asexual fruiting body of Dictyostelium discoideum using haploid and isogenic diploid strains.Differentiation. 1981;18(1):1-9. doi: 10.1111/j.1432-0436.1981.tb01099.x. Differentiation. 1981. PMID: 7227705
-
Cancer and the breakdown of multicellularity: What Dictyostelium discoideum, a social amoeba, can teach us.Bioessays. 2021 Apr;43(4):e2000156. doi: 10.1002/bies.202000156. Epub 2021 Jan 14. Bioessays. 2021. PMID: 33448043 Review.
-
Regulation of cell-fate determination in Dictyostelium.Dev Biol. 1999 Dec 15;216(2):426-41. doi: 10.1006/dbio.1999.9485. Dev Biol. 1999. PMID: 10642783 Review.
-
Prestalk-like positioning of de-differentiated cells in the social amoeba Dictyostelium discoideum.Sci Rep. 2024 Apr 1;14(1):7677. doi: 10.1038/s41598-024-58277-3. Sci Rep. 2024. PMID: 38561423 Free PMC article.
-
Inhibition of multicellular development switches cell death of Dictyostelium discoideum towards mammalian-like unicellular apoptosis.Eur J Cell Biol. 2001 Jun;80(6):428-41. doi: 10.1078/0171-9335-00172. Eur J Cell Biol. 2001. PMID: 11484934
Cited by
-
Relatedness and the evolution of mechanisms to divide labor in microorganisms.Ecol Evol. 2021 Oct 8;11(21):14475-14489. doi: 10.1002/ece3.8067. eCollection 2021 Nov. Ecol Evol. 2021. PMID: 34765120 Free PMC article.
-
Life cycle strategies in free-living unicellular eukaryotes: Diversity, evolution, and current molecular tools to unravel the private life of microorganisms.J Eukaryot Microbiol. 2024 Nov-Dec;71(6):e13052. doi: 10.1111/jeu.13052. Epub 2024 Jul 31. J Eukaryot Microbiol. 2024. PMID: 39085163 Free PMC article. Review.
References
-
- Aerts R. J., Durston A. J., Konijn T. M. (1987). Cytoplasmic pH at the onset of development in Dictyostelium. J. Cell Sci. 87 423–430. - PubMed
-
- Ambros V. (1999). Cell cycle-dependent sequencing of cell fate decisions in Caenorhabditis elegans vulva precursor cells. Development 126 1947–1956. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials