

Transcriptomics of Meiosis in the Male Mouse
- PMID: 33748111
- PMCID: PMC7973102
- DOI: 10.3389/fcell.2021.626020
Transcriptomics of Meiosis in the Male Mouse
Abstract
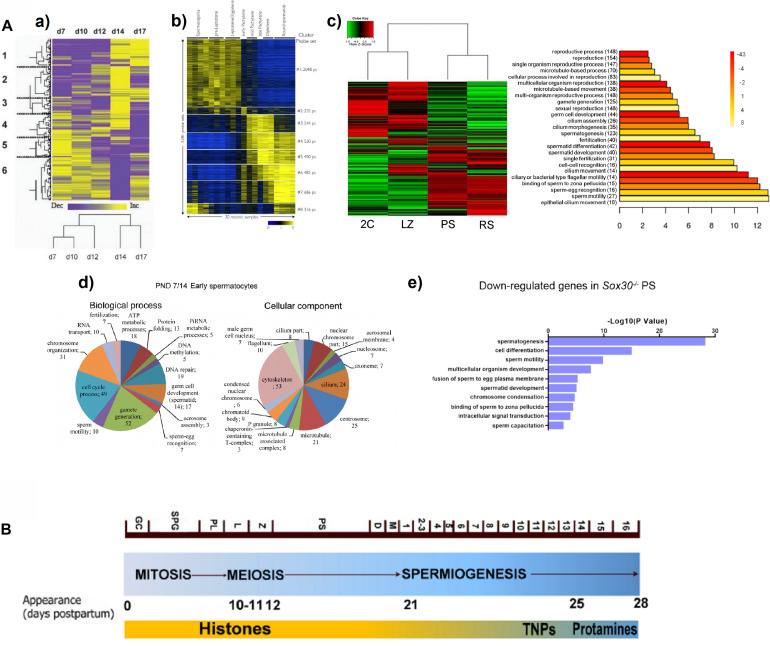
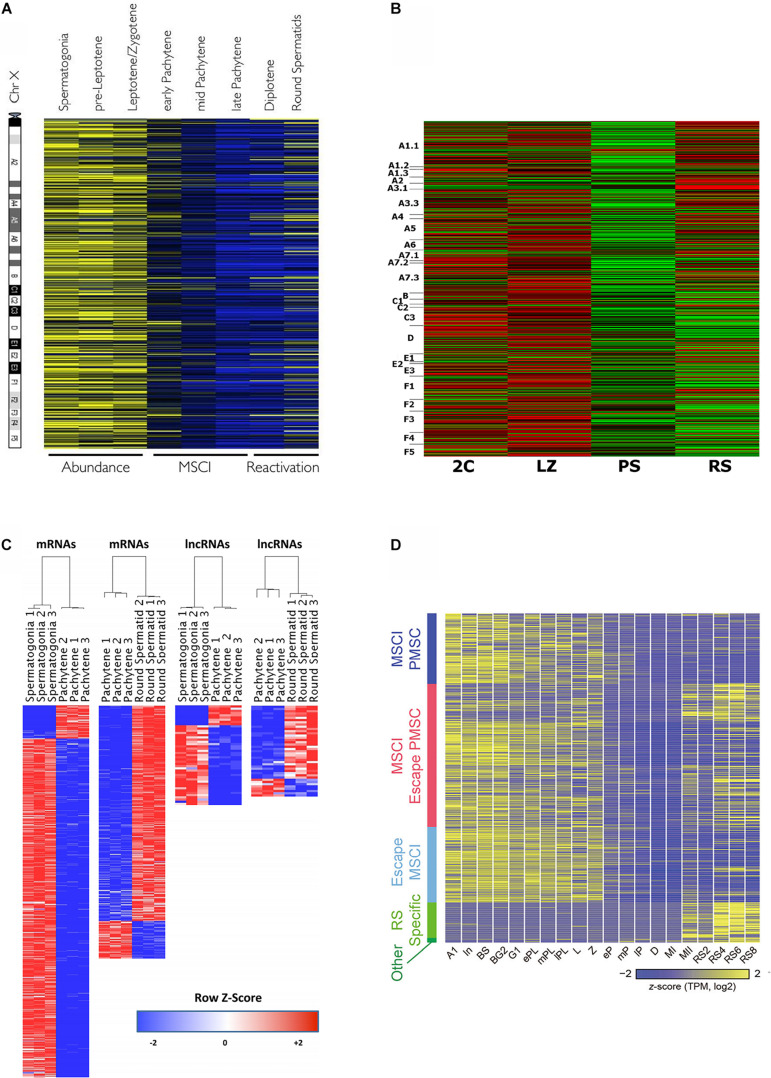
Molecular studies of meiosis in mammals have been long relegated due to some intrinsic obstacles, namely the impossibility to reproduce the process in vitro, and the difficulty to obtain highly pure isolated cells of the different meiotic stages. In the recent years, some technical advances, from the improvement of flow cytometry sorting protocols to single-cell RNAseq, are enabling to profile the transcriptome and its fluctuations along the meiotic process. In this mini-review we will outline the diverse methodological approaches that have been employed, and some of the main findings that have started to arise from these studies. As for practical reasons most studies have been carried out in males, and mostly using mouse as a model, our focus will be on murine male meiosis, although also including specific comments about humans. Particularly, we will center on the controversy about gene expression during early meiotic prophase; the widespread existing gap between transcription and translation in meiotic cells; the expression patterns and potential roles of meiotic long non-coding RNAs; and the visualization of meiotic sex chromosome inactivation from the RNAseq perspective.
Keywords: MSCI; RNAseq; lncRNAs; meiosis; meiotic prophase; spermatogenesis; spermatogenic cell sorting; transcriptomics.
Copyright © 2021 Geisinger, Rodríguez-Casuriaga and Benavente.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous