

The membrane axis of Alzheimer's nanomedicine
- PMID: 33748816
- PMCID: PMC7971452
- DOI: 10.1002/anbr.202000040
The membrane axis of Alzheimer's nanomedicine
Abstract
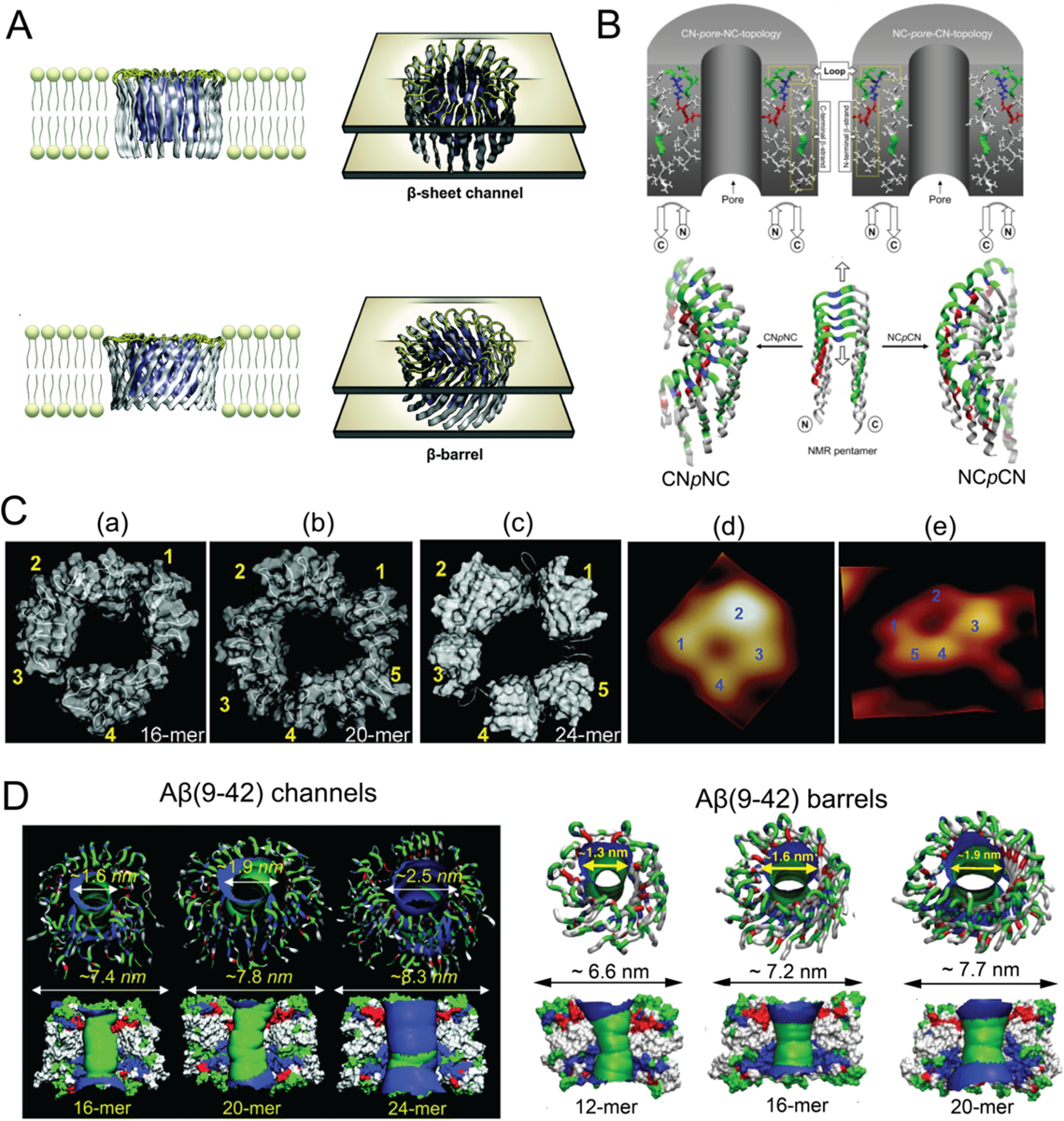
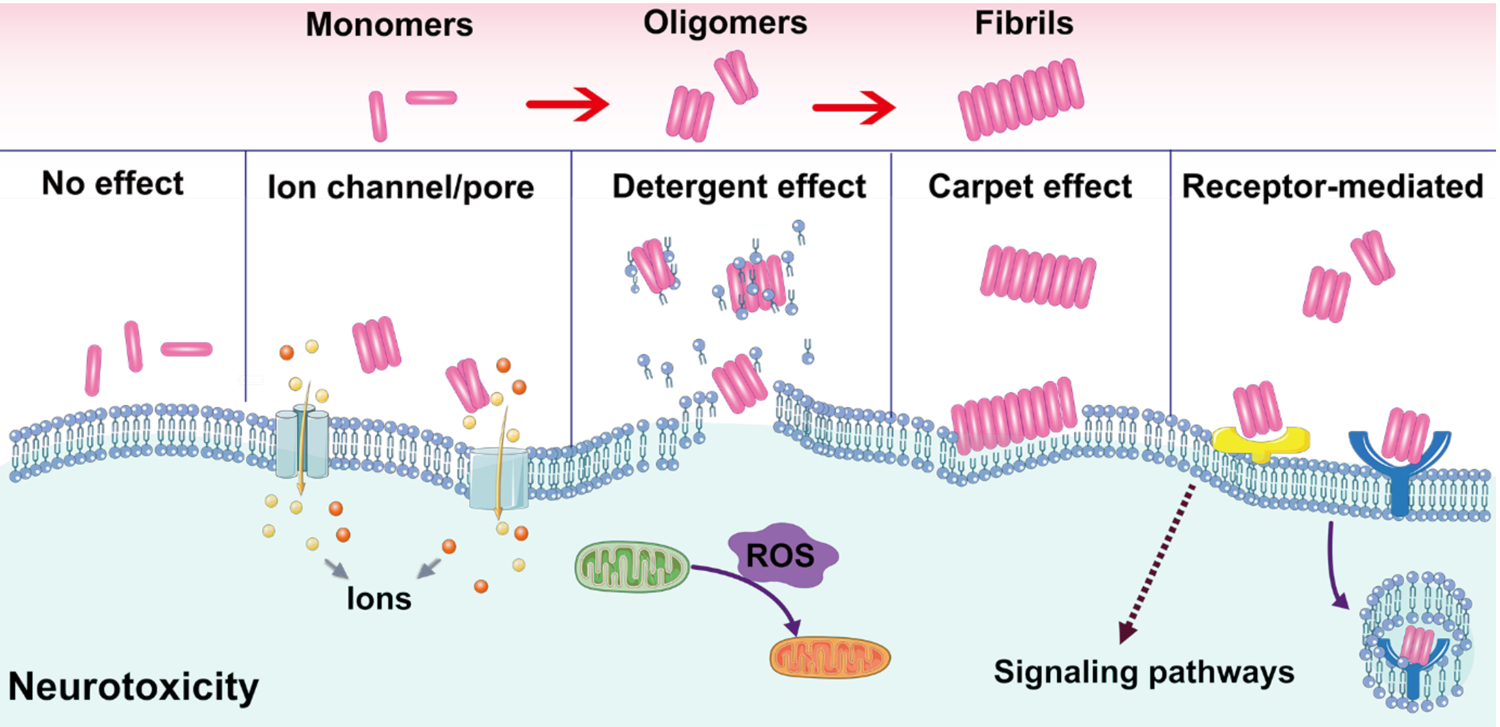
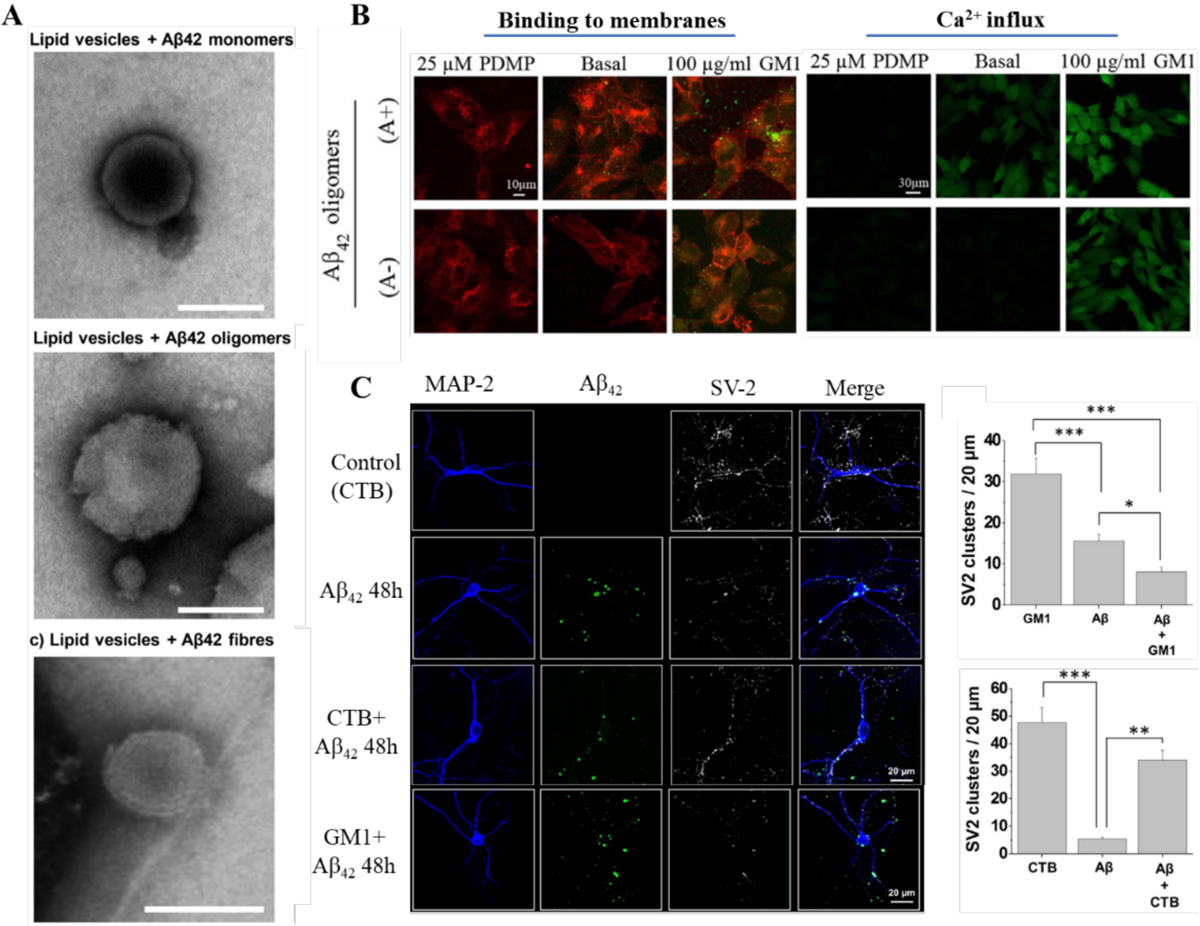
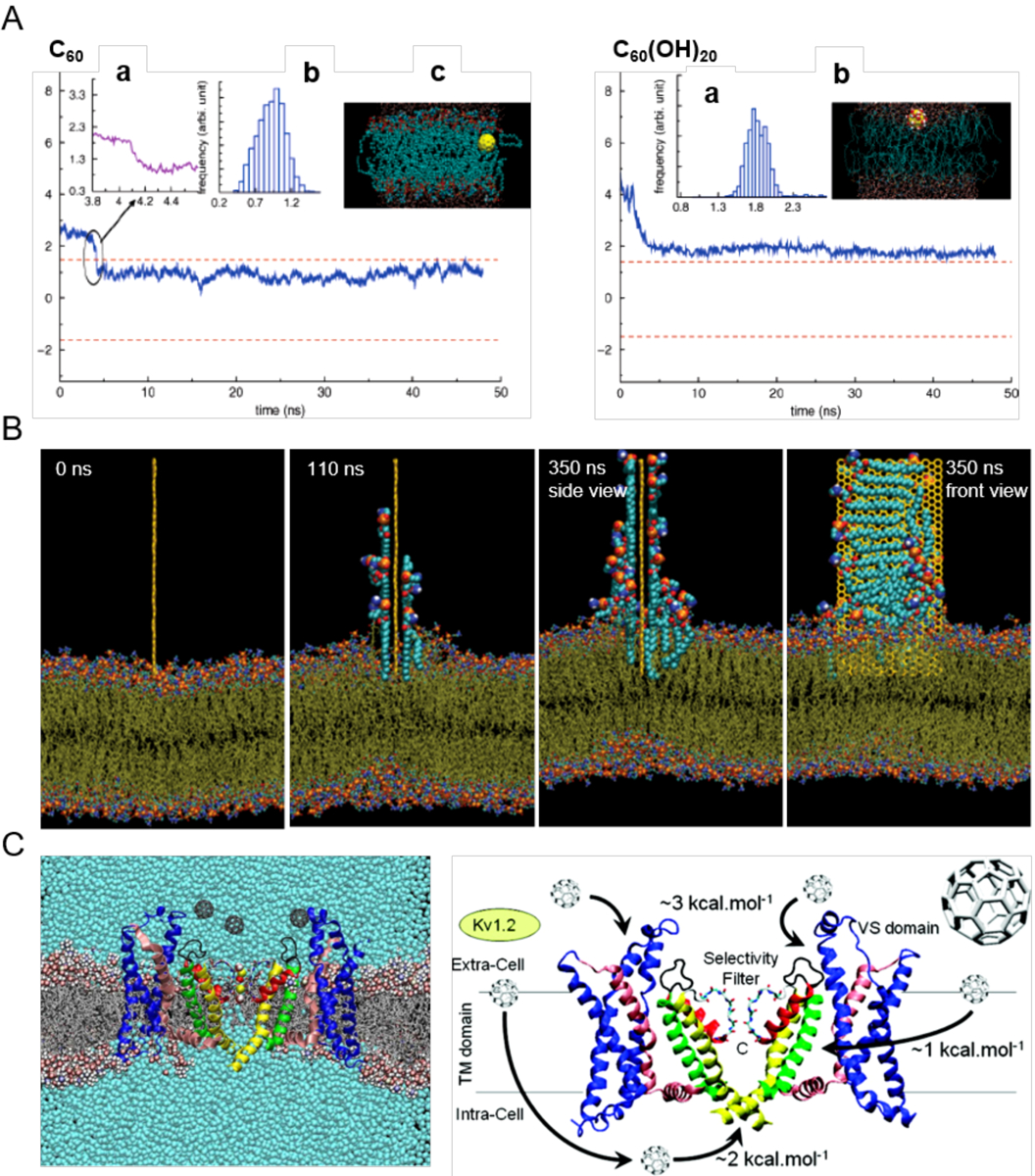
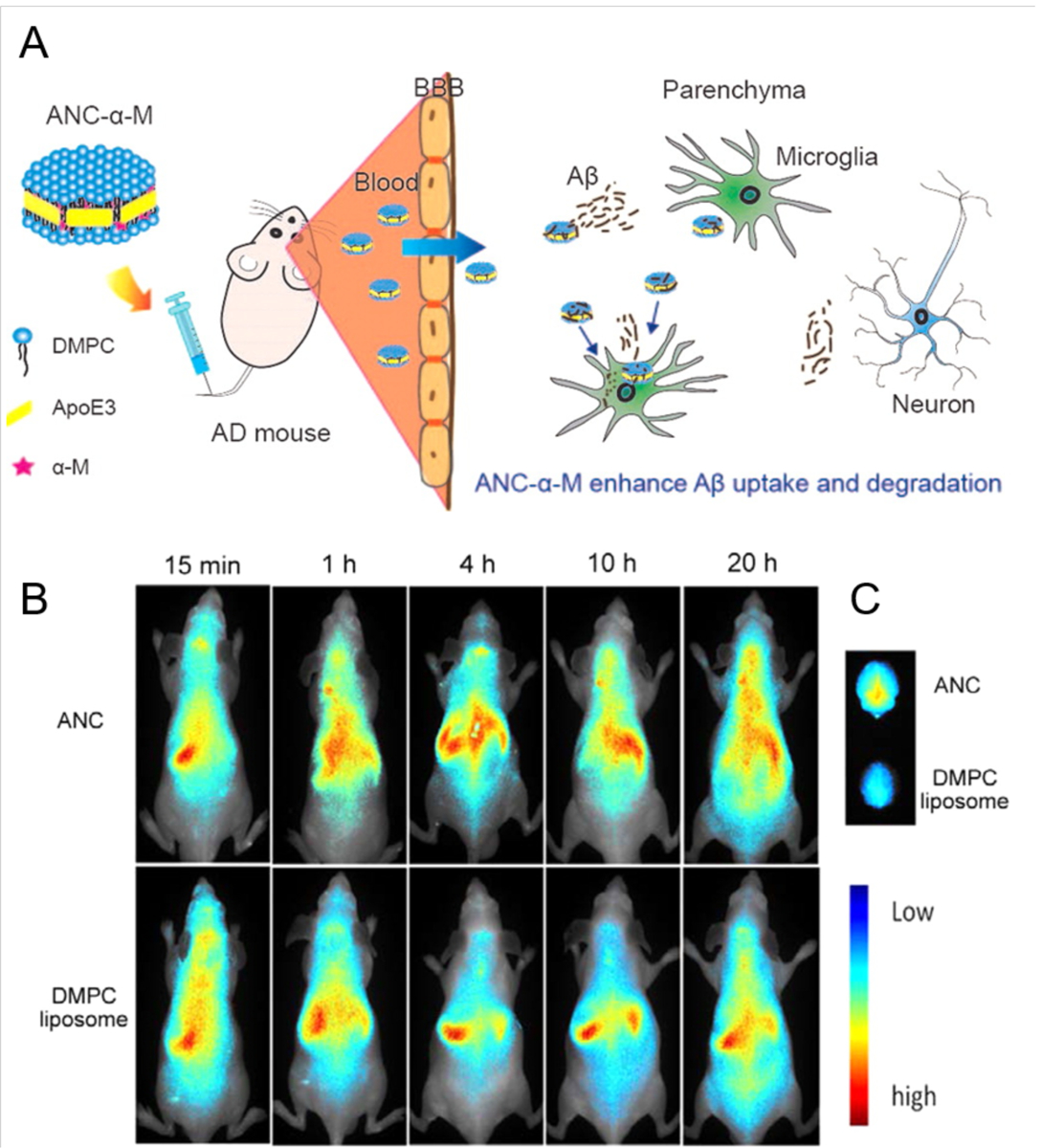
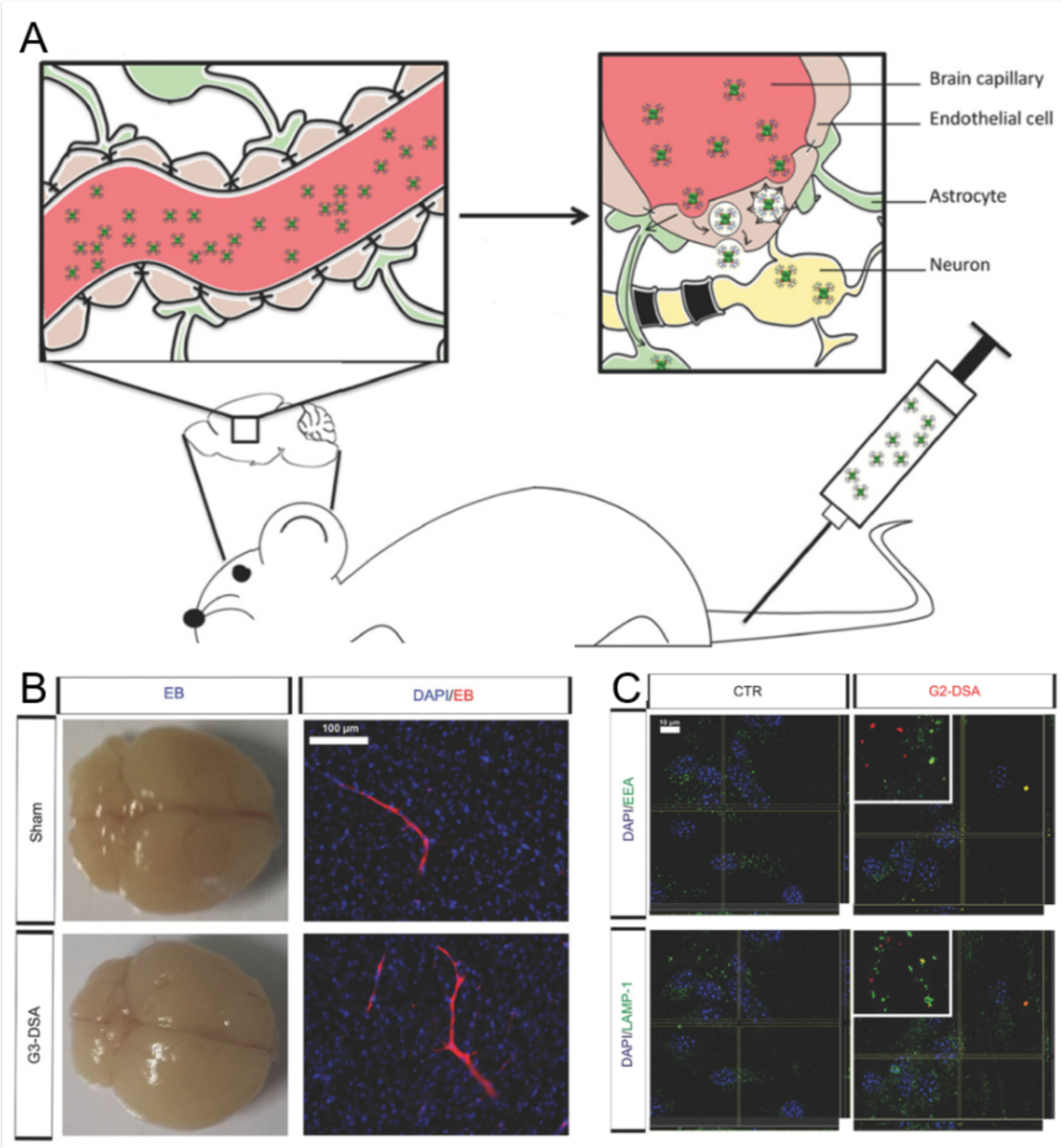
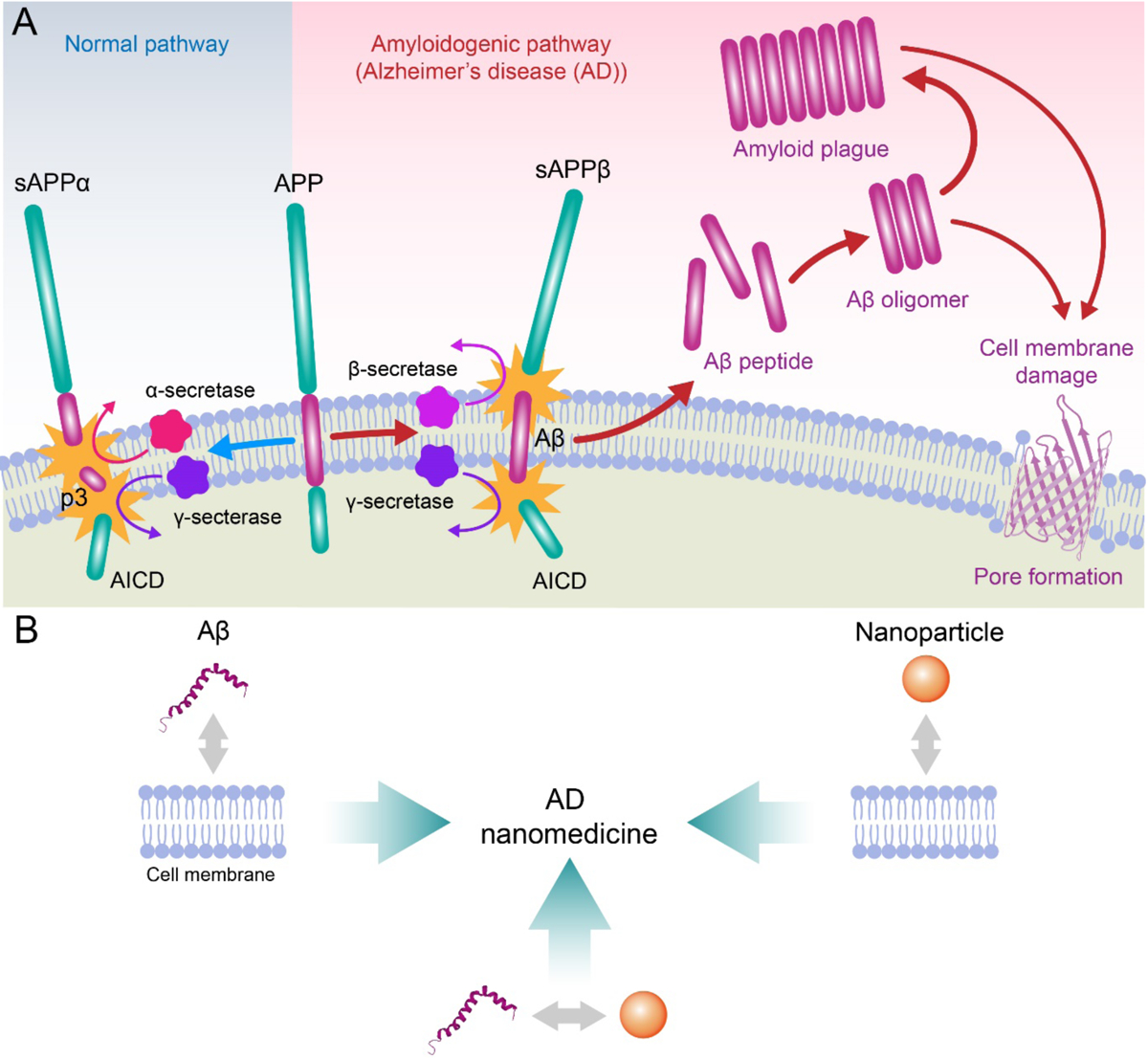
Alzheimer's disease (AD) is a major neurological disorder impairing its carrier's cognitive function, memory and lifespan. While the development of AD nanomedicine is still nascent, the field is evolving into a new scientific frontier driven by the diverse physicochemical properties and theranostic potential of nanomaterials and nanocomposites. Characteristic to the AD pathology is the deposition of amyloid plaques and tangles of amyloid beta (Aβ) and tau, whose aggregation kinetics may be curbed by nanoparticle inhibitors via sequence-specific targeting or nonspecific interactions with the amyloidogenic proteins. As literature implicates cell membrane as a culprit in AD pathogenesis, here we summarize the membrane axis of AD nanomedicine and present a new rationale that the field development may greatly benefit from harnessing our existing knowledge of Aβ-membrane interaction, nanoparticle-membrane interaction and Aβ-nanoparticle interaction.
Keywords: AD; Aβ; membrane; nanomedicine; nanoparticle.
Conflict of interest statement
Declaration of Competing Interest There are no conflicts to declare.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials