

Amplification of potential thermogenetic mechanisms in cetacean brains compared to artiodactyl brains
- PMID: 33750832
- PMCID: PMC7970898
- DOI: 10.1038/s41598-021-84762-0
Amplification of potential thermogenetic mechanisms in cetacean brains compared to artiodactyl brains
Abstract
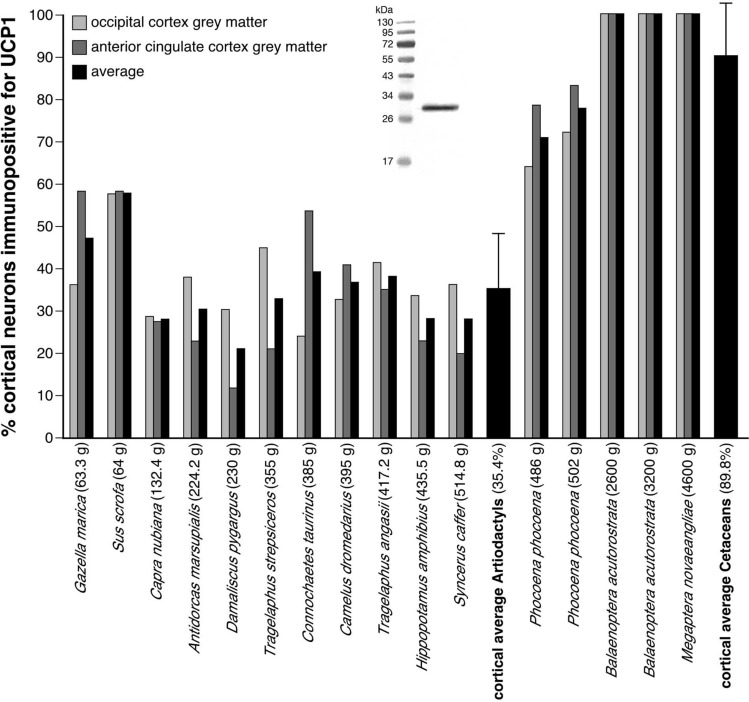
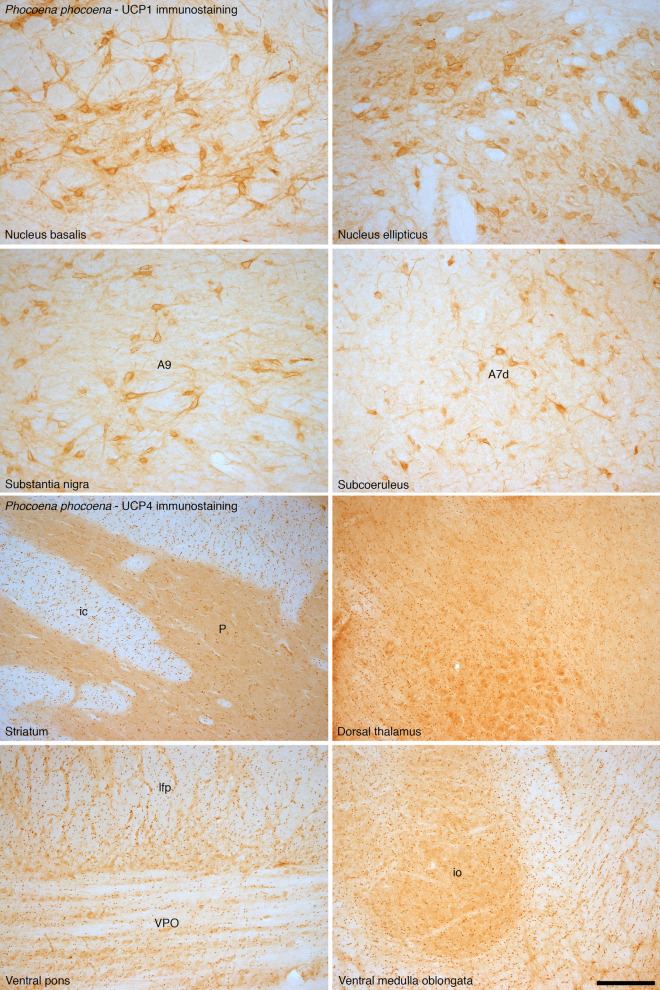
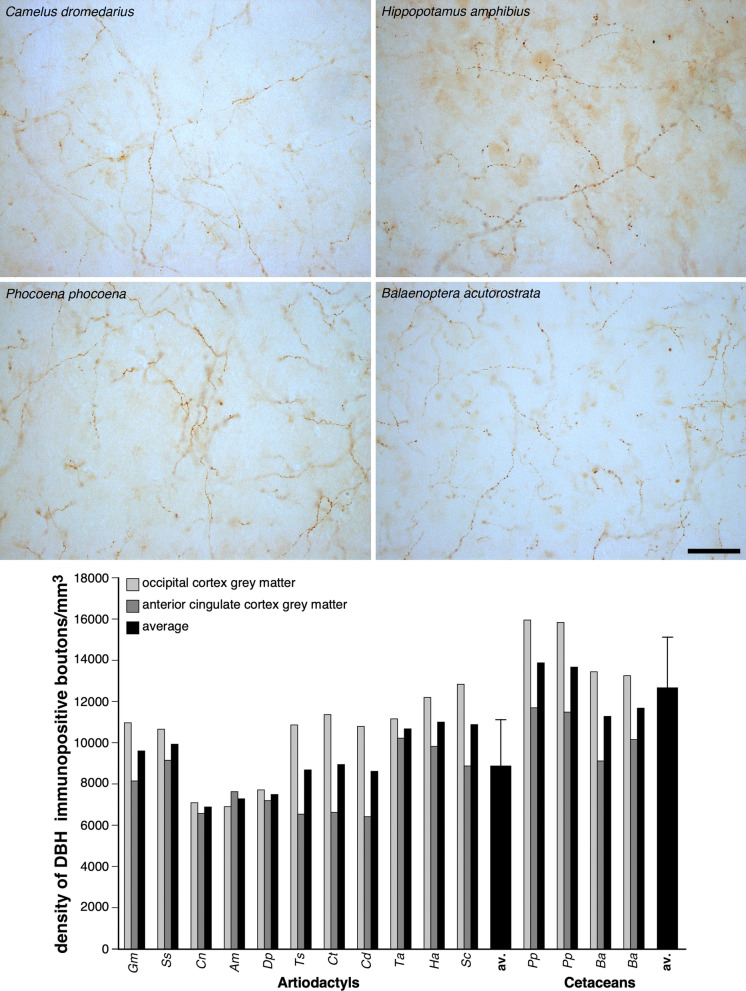
To elucidate factors underlying the evolution of large brains in cetaceans, we examined 16 brains from 14 cetartiodactyl species, with immunohistochemical techniques, for evidence of non-shivering thermogenesis. We show that, in comparison to the 11 artiodactyl brains studied (from 11 species), the 5 cetacean brains (from 3 species), exhibit an expanded expression of uncoupling protein 1 (UCP1, UCPs being mitochondrial inner membrane proteins that dissipate the proton gradient to generate heat) in cortical neurons, immunolocalization of UCP4 within a substantial proportion of glia throughout the brain, and an increased density of noradrenergic axonal boutons (noradrenaline functioning to control concentrations of and activate UCPs). Thus, cetacean brains studied possess multiple characteristics indicative of intensified thermogenetic functionality that can be related to their current and historical obligatory aquatic niche. These findings necessitate reassessment of our concepts regarding the reasons for large brain evolution and associated functional capacities in cetaceans.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Marino L, et al. A claim in search of evidence: reply to Manger’s thermogenesis hypothesis of cetacean brain structure. Biol. Rev. 2008;83:417–440. - PubMed
-
- Kesarev VS. The inferior brain of the dolphin. Soviet Sci. Rev. 1971;1:52–58.
-
- Nikolskaya KA. Evolutionary aspects of intellect in vertebrates: Can intellect be a factor confining choice of the habitat? Invest. Russ. 2005;8:1442–1500.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
