

Deconstructing the contribution of sensory cues in social approach
- PMID: 33751673
- PMCID: PMC8251867
- DOI: 10.1111/ejn.15179
Deconstructing the contribution of sensory cues in social approach
Abstract
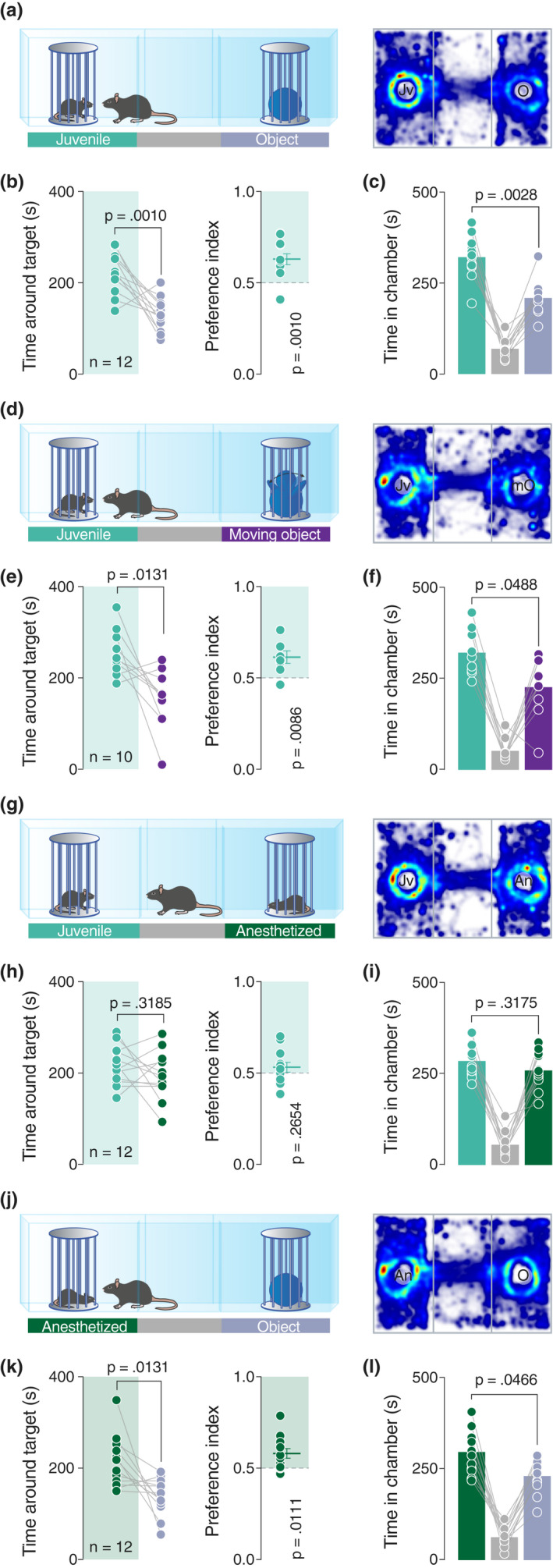
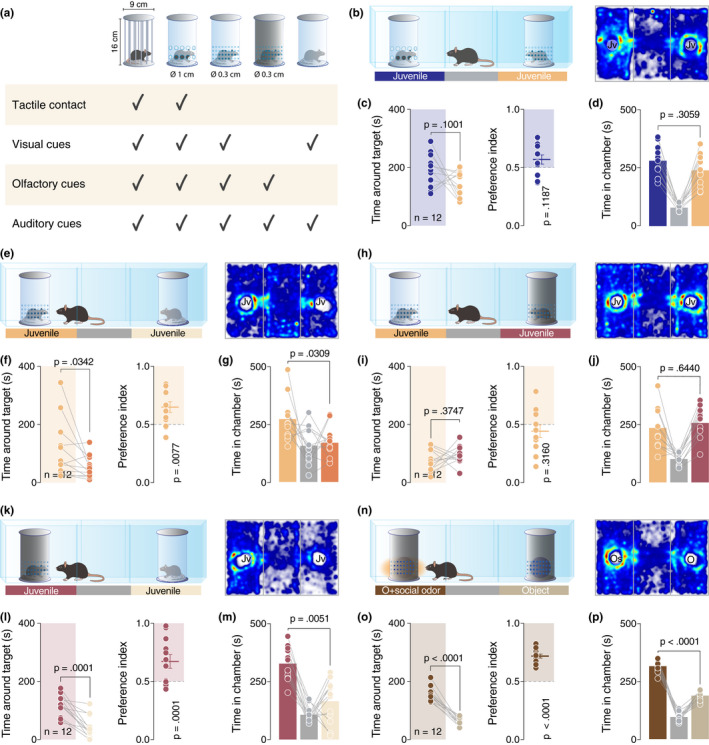
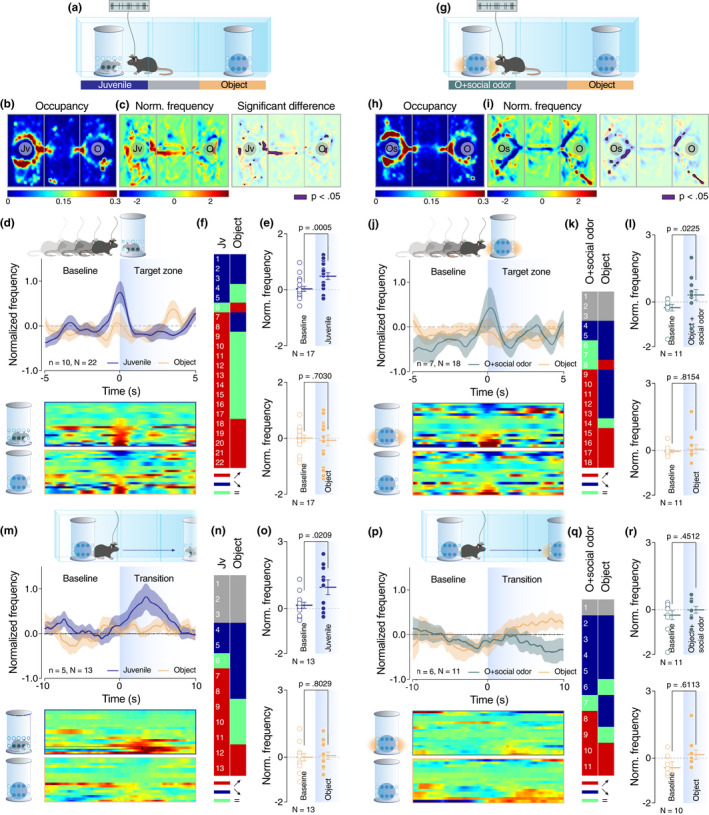
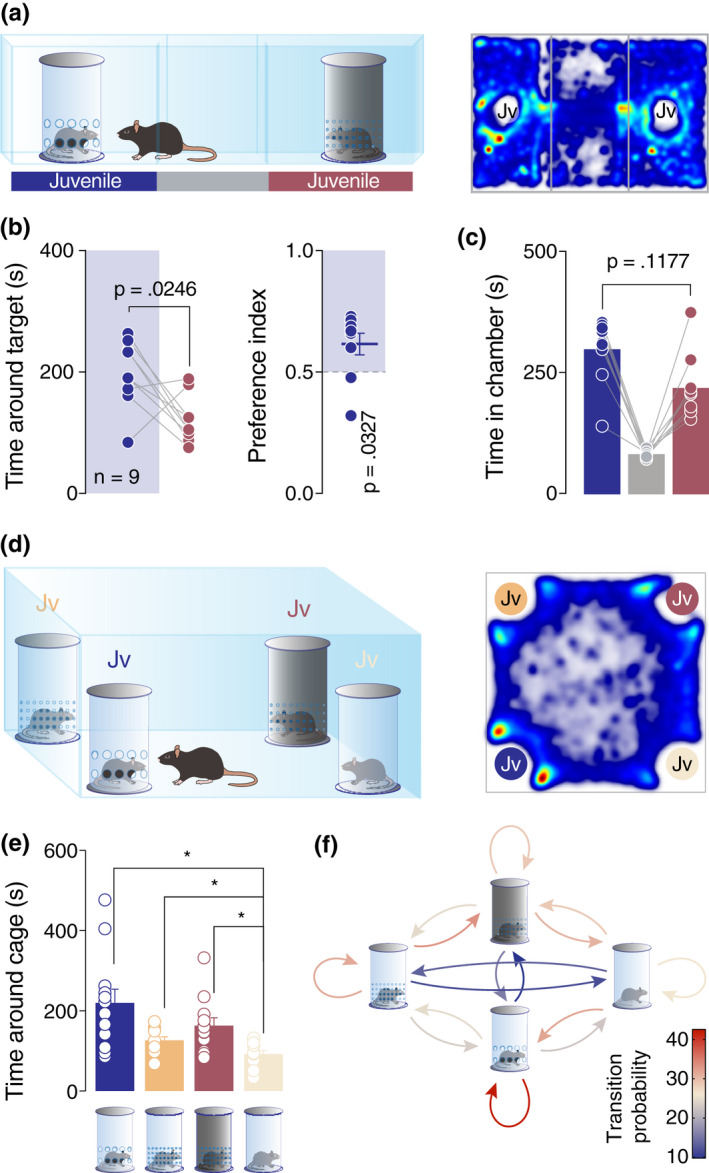
Social interaction is a complex and highly conserved behavior that safeguards survival and reproductive success. Although considerable progress has been made regarding our understanding of same-sex conspecific and non-aggressive interactions, questions regarding the precise contribution of sensory cues in social approach and their specific neurobiological correlates remain open. Here, by designing a series of experiments with diverse social and object stimuli manipulations in custom-made enclosures, we first sought to deconstruct key elements of social preference as assessed by the three-chamber task. Our results highlight the importance of social olfactory cues in approach behavior. Subsequently, we interrogated whether a social odor would activate dopaminergic neurons of the Ventral Tegmental Area in the same way as a juvenile conspecific would. Employing in vivo recordings in freely behaving mice, we observed an increase of the firing only during the transition toward the juvenile mouse and not during the transition toward the object impregnated with social odor, suggesting that these two experiences are distinct and can be differentiated at the neuronal level. Moreover, using a four-choice task, we further showed that mice prefer to explore complex social stimuli compared to isolated sensory cues. Our findings offer insights toward understanding how different sensory modalities contribute to the neurobiological basis of social behavior which can be essential when studying social deficits observed in autism-, depression-, anxiety-, or schizophrenia-related mouse models.
Keywords: VTA; dopamine; sensory cues; sociability; three-chamber test.
© 2021 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bariselli, S. , Contestabile, A. , Tzanoulinou, S. , Musardo, S. , & Bellone, C. (2018). SHANK3 downregulation in the ventral tegmental area accelerates the extinction of contextual associations induced by juvenile non‐familiar conspecific interaction. Frontiers in Molecular Neuroscience, 11, 360. 10.3389/fnmol.2018.00360 - DOI - PMC - PubMed
-
- Bariselli, S. , Hörnberg, H. , Prévost‐Solié, C. , Musardo, S. , Hatstatt‐Burklé, L. , Scheiffele, P. , & Bellone, C. (2018). Role of VTA dopamine neurons and neuroligin 3 in sociability traits related to nonfamiliar conspecific interaction. Nature Communications, 9, 3173. 10.1038/s41467-018-05382-3 - DOI - PMC - PubMed
-
- Bariselli, S. , Tzanoulinou, S. , Glangetas, C. , Prévost‐Solié, C. , Pucci, L. , Viguié, J. , Bezzi, P. , O'Connor, E. C. , Georges, F. , Lüscher, C. , & Bellone, C. (2016). SHANK3 controls maturation of social reward circuits in the VTA. Nature Neuroscience, 19, 926–934. 10.1038/nn.4319 - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
