

The development of adult intestinal stem cells: Insights from studies on thyroid hormone-dependent anuran metamorphosis
- PMID: 33752821
- PMCID: PMC12268621
- DOI: 10.1016/bs.vh.2021.02.010
The development of adult intestinal stem cells: Insights from studies on thyroid hormone-dependent anuran metamorphosis
Abstract
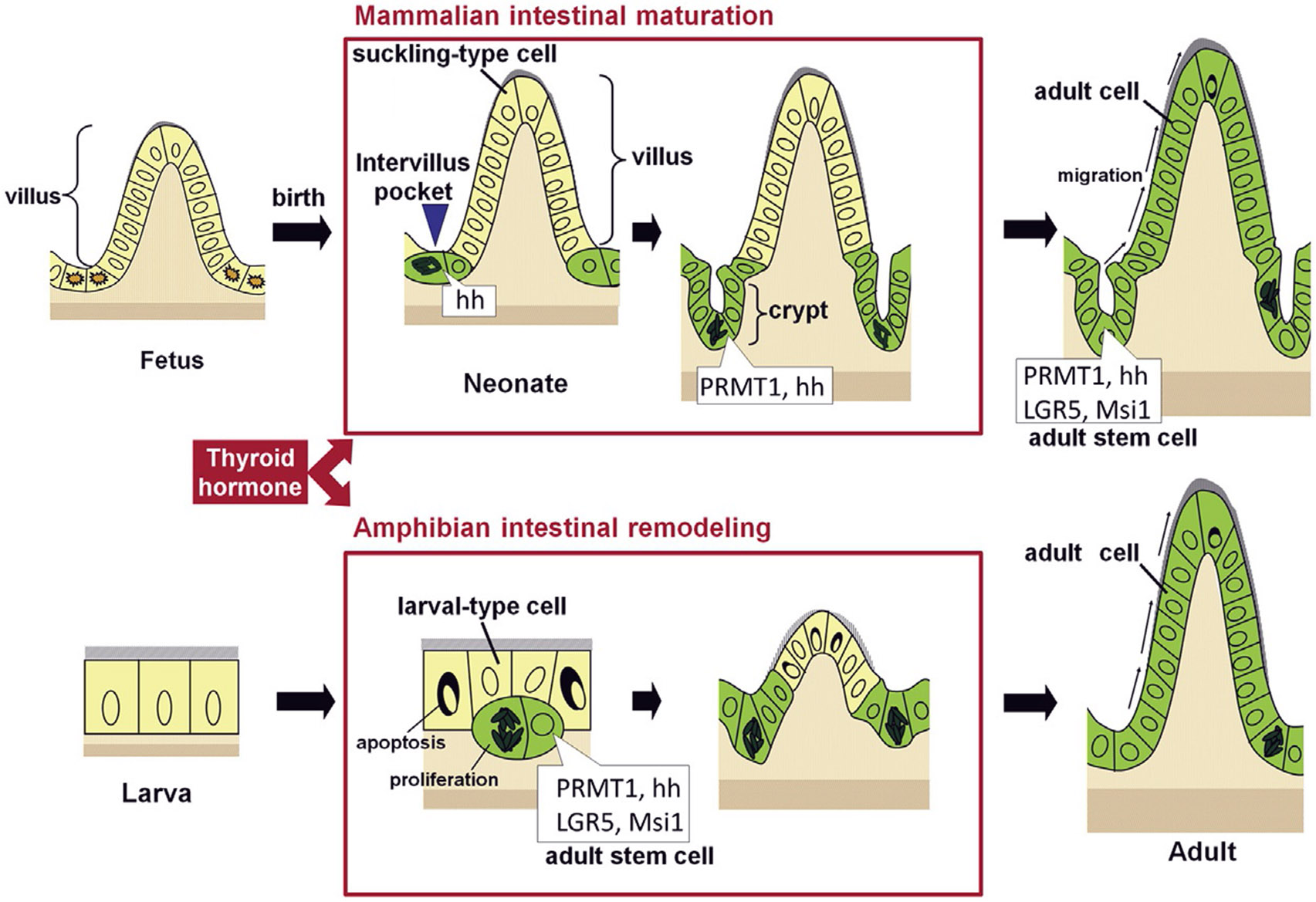
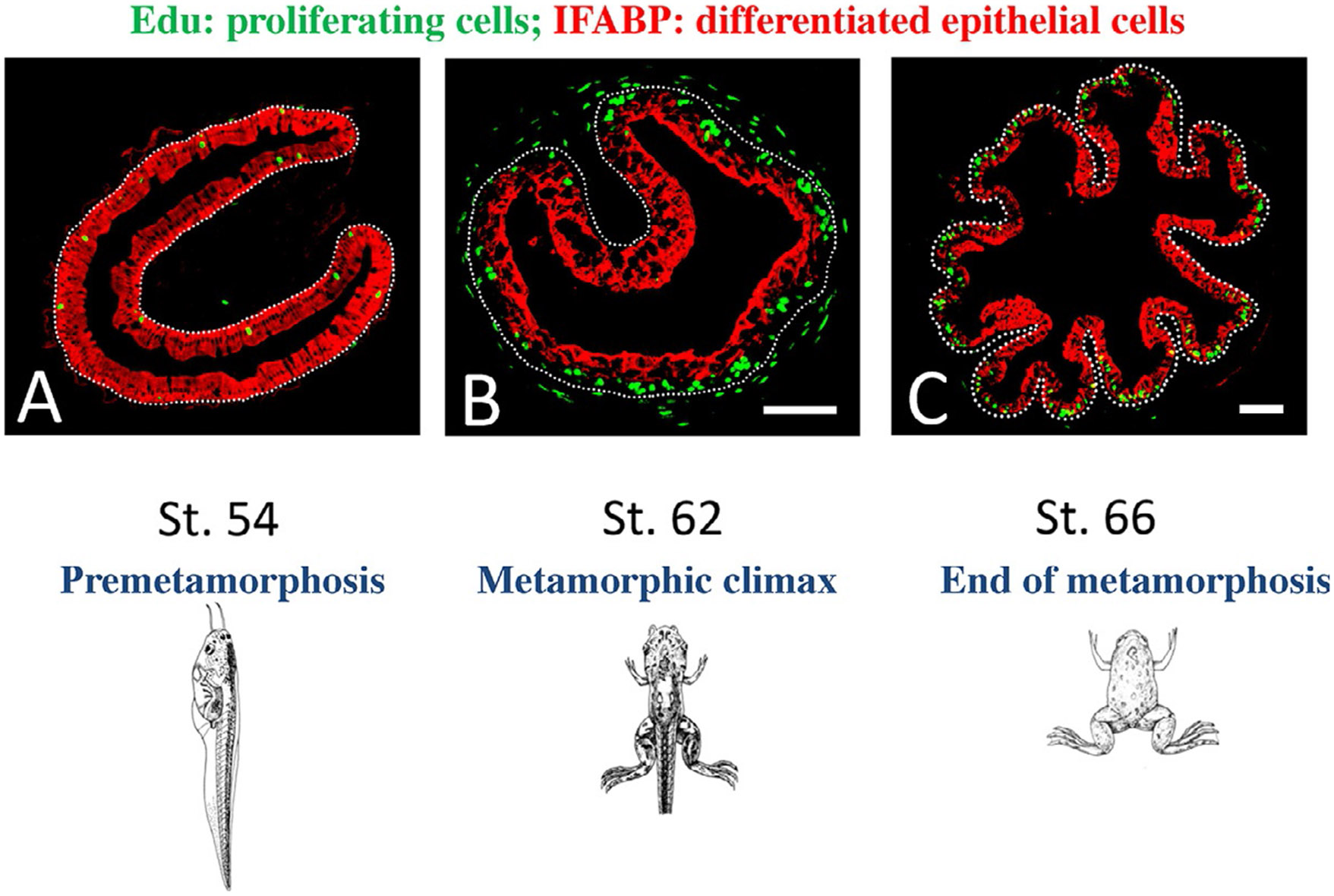
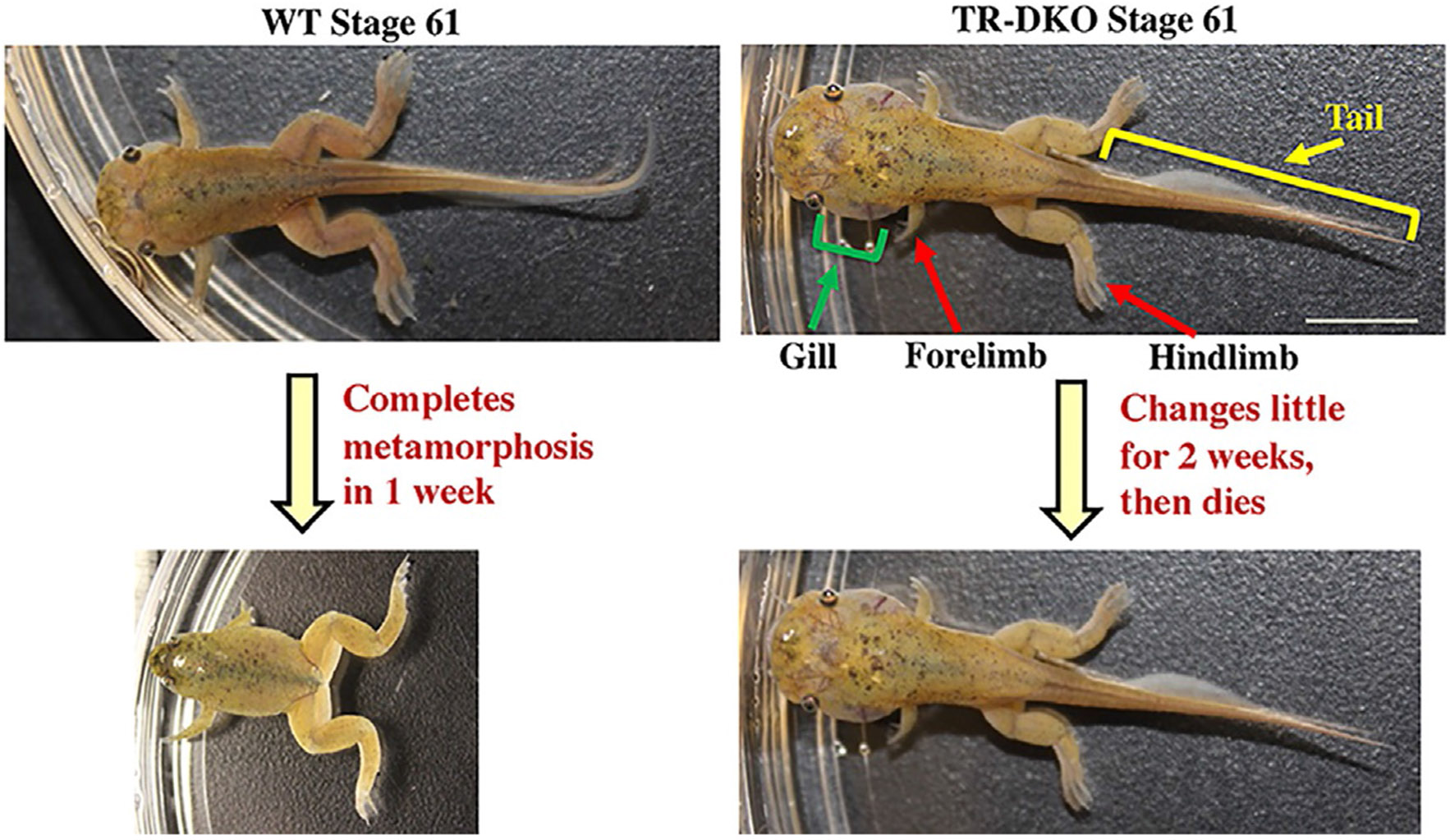
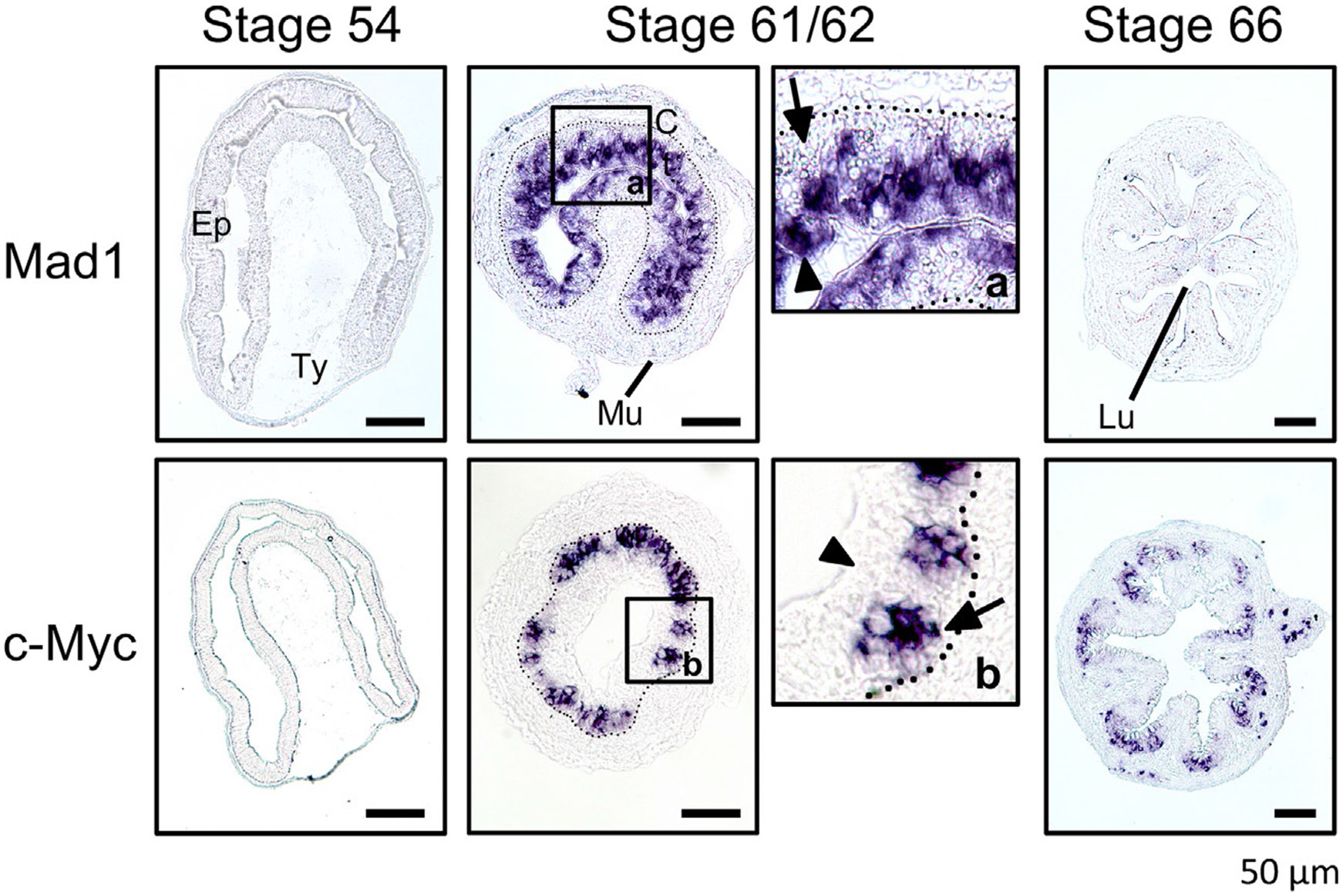
Vertebrates organ development often takes place in two phases: initial formation and subsequent maturation into the adult form. This is exemplified by the intestine. In mouse, the intestine at birth has villus, where most differentiated epithelial cells are located, but lacks any crypts, where adult intestinal stem cells reside. The crypt is formed during the first 3 weeks after birth when plasma thyroid hormone (T3) levels are high. Similarly, in anurans, the intestine undergoes drastic remodeling into the adult form during metamorphosis in a process completely dependent on T3. Studies on Xenopus metamorphosis have revealed important clues on the formation of the adult intestine during metamorphosis. Here we will review our current understanding on how T3 induces the degeneration of larval epithelium and de novo formation of adult intestinal stem cells. We will also discuss the mechanistic conservations in intestinal development between anurans and mammals.
Keywords: Intestine; Metamorphosis; Postembryonic development; Stem cell; Thyroid hormone receptor; Xenopus laevis; Xenopus tropicalis.
Copyright © 2021 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Intestinal remodeling during Xenopus metamorphosis as a model for studying thyroid hormone signaling and adult organogenesis.Mol Cell Endocrinol. 2024 May 15;586:112193. doi: 10.1016/j.mce.2024.112193. Epub 2024 Feb 22. Mol Cell Endocrinol. 2024. PMID: 38401883 Free PMC article.
-
Thyroid hormone directly activates mitochondrial fission process 1 (Mtfp1) gene transcription during adult intestinal stem cell development and proliferation in Xenopus tropicalis.Gen Comp Endocrinol. 2020 Dec 1;299:113590. doi: 10.1016/j.ygcen.2020.113590. Epub 2020 Aug 20. Gen Comp Endocrinol. 2020. PMID: 32827515 Free PMC article.
-
Organ-specific effects on target binding due to knockout of thyroid hormone receptor α during Xenopus metamorphosis.Dev Growth Differ. 2023 Jan;65(1):23-28. doi: 10.1111/dgd.12825. Epub 2022 Dec 1. Dev Growth Differ. 2023. PMID: 36397722 Free PMC article.
-
Thyroid hormone receptor subtype-specific function in controlling organ-specific developmental timing and rate during Xenopus development.Front Endocrinol (Lausanne). 2025 Jun 9;16:1614439. doi: 10.3389/fendo.2025.1614439. eCollection 2025. Front Endocrinol (Lausanne). 2025. PMID: 40551888 Free PMC article. Review.
-
Thyroid hormone regulation of adult intestinal stem cell development: mechanisms and evolutionary conservations.Int J Biol Sci. 2012;8(8):1217-24. doi: 10.7150/ijbs.5109. Epub 2012 Oct 23. Int J Biol Sci. 2012. PMID: 23136549 Free PMC article. Review.
Cited by
-
Effects of hormones on intestinal stem cells.Stem Cell Res Ther. 2023 Apr 26;14(1):105. doi: 10.1186/s13287-023-03336-1. Stem Cell Res Ther. 2023. PMID: 37101229 Free PMC article. Review.
-
Liver development during Xenopus tropicalis metamorphosis is controlled by T3-activation of WNT signaling.iScience. 2023 Feb 28;26(4):106301. doi: 10.1016/j.isci.2023.106301. eCollection 2023 Apr 21. iScience. 2023. PMID: 37153451 Free PMC article.
-
Regulation of the THRA gene, encoding the thyroid hormone nuclear receptor TRα1, in intestinal lesions.Mol Oncol. 2022 Dec;16(22):3975-3993. doi: 10.1002/1878-0261.13298. Epub 2022 Oct 10. Mol Oncol. 2022. PMID: 36217307 Free PMC article.
-
Intestinal remodeling during Xenopus metamorphosis as a model for studying thyroid hormone signaling and adult organogenesis.Mol Cell Endocrinol. 2024 May 15;586:112193. doi: 10.1016/j.mce.2024.112193. Epub 2024 Feb 22. Mol Cell Endocrinol. 2024. PMID: 38401883 Free PMC article.
-
Mucosal Immunity and the Gut-Microbiota-Brain-Axis in Neuroimmune Disease.Int J Mol Sci. 2022 Nov 1;23(21):13328. doi: 10.3390/ijms232113328. Int J Mol Sci. 2022. PMID: 36362150 Free PMC article. Review.
References
-
- Amano T, & Yoshizato K (1998). Isolation of genes involved in intestinal remodeling during anuran metamorphosis. Wound Repair and Regeneration, 6, 302–313. - PubMed
-
- Amati B, & Land H (1994). Myc-Max-Mad: A transcription factor network controlling cell cycle progression, differentiation and death. Current Opinion in Genetics & Development, 4, 102–108. - PubMed
-
- Bagamasbad P, Howdeshell KL, Sachs LM, Demeneix BA, & Denver RJ (2008). A role for basic transcription element-binding protein 1 (BTEB1) in the autoinduction of thyroid hormone receptor beta. The Journal of Biological Chemistry, 283, 2275–2285. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
