

Mitochondrial metabolism is essential for invariant natural killer T cell development and function
- PMID: 33753493
- PMCID: PMC8020658
- DOI: 10.1073/pnas.2021385118
Mitochondrial metabolism is essential for invariant natural killer T cell development and function
Abstract
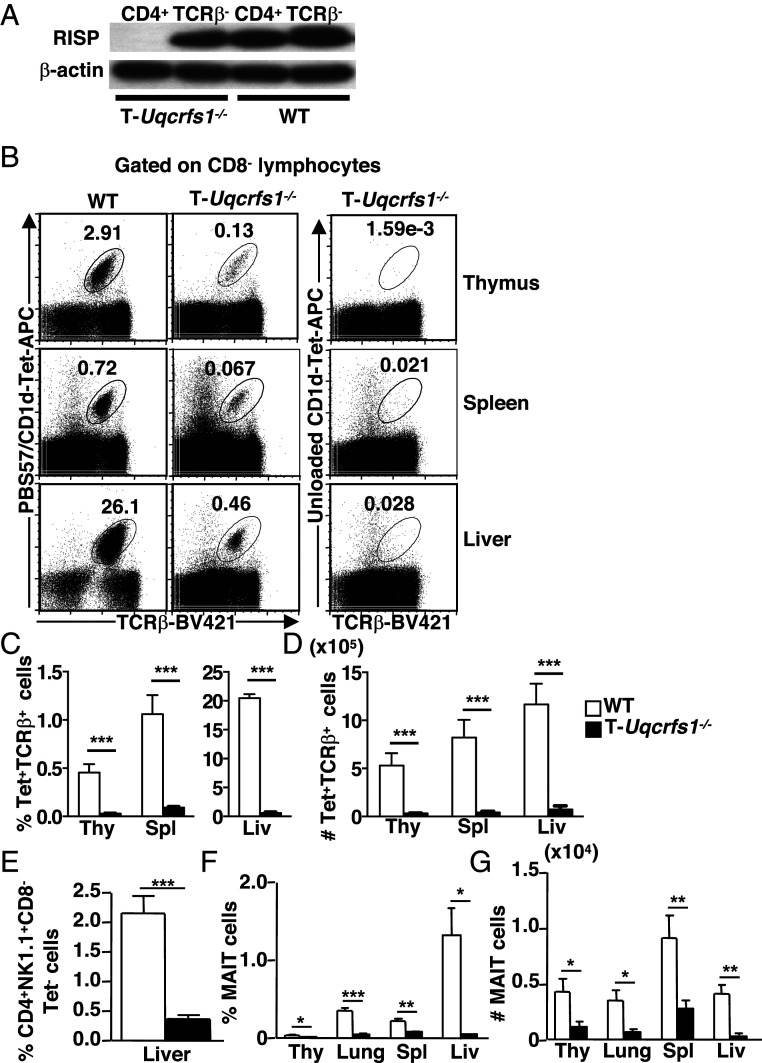
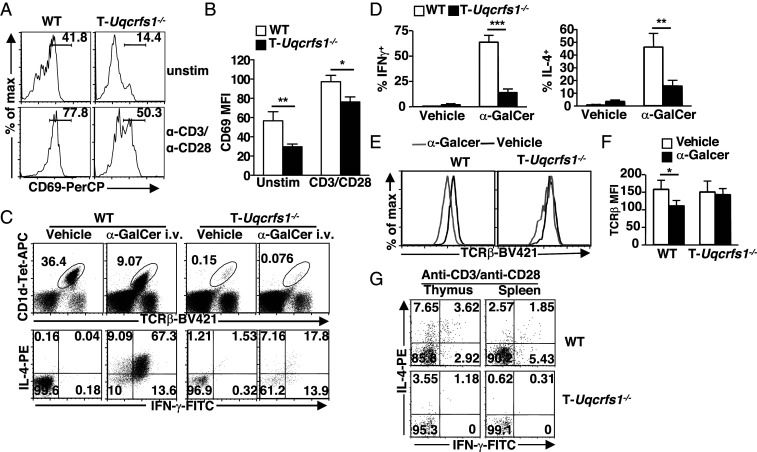
Conventional T cell fate and function are determined by coordination between cellular signaling and mitochondrial metabolism. Invariant natural killer T (iNKT) cells are an important subset of "innate-like" T cells that exist in a preactivated effector state, and their dependence on mitochondrial metabolism has not been previously defined genetically or in vivo. Here, we show that mature iNKT cells have reduced mitochondrial respiratory reserve and iNKT cell development was highly sensitive to perturbation of mitochondrial function. Mice with T cell-specific ablation of Rieske iron-sulfur protein (RISP; T-Uqcrfs1-/- ), an essential subunit of mitochondrial complex III, had a dramatic reduction of iNKT cells in the thymus and periphery, but no significant perturbation on the development of conventional T cells. The impaired development observed in T-Uqcrfs1-/- mice stems from a cell-autonomous defect in iNKT cells, resulting in a differentiation block at the early stages of iNKT cell development. Residual iNKT cells in T-Uqcrfs1-/- mice displayed increased apoptosis but retained the ability to proliferate in vivo, suggesting that their bioenergetic and biosynthetic demands were not compromised. However, they exhibited reduced expression of activation markers, decreased T cell receptor (TCR) signaling and impaired responses to TCR and interleukin-15 stimulation. Furthermore, knocking down RISP in mature iNKT cells diminished their cytokine production, correlating with reduced NFATc2 activity. Collectively, our data provide evidence for a critical role of mitochondrial metabolism in iNKT cell development and activation outside of its traditional role in supporting cellular bioenergetic demands.
Keywords: CD1; NKT cells; T cell development; knockout mice; mitochondrial metabolism.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures






Similar articles
-
Essential role for autophagy during invariant NKT cell development.Proc Natl Acad Sci U S A. 2014 Dec 30;111(52):E5678-87. doi: 10.1073/pnas.1413935112. Epub 2014 Dec 15. Proc Natl Acad Sci U S A. 2014. PMID: 25512546 Free PMC article.
-
Discrete TCR Binding Kinetics Control Invariant NKT Cell Selection and Central Priming.J Immunol. 2016 Nov 15;197(10):3959-3969. doi: 10.4049/jimmunol.1601382. Epub 2016 Oct 19. J Immunol. 2016. PMID: 27798168
-
New Genetically Manipulated Mice Provide Insights Into the Development and Physiological Functions of Invariant Natural Killer T Cells.Front Immunol. 2018 Jun 14;9:1294. doi: 10.3389/fimmu.2018.01294. eCollection 2018. Front Immunol. 2018. PMID: 29963043 Free PMC article. Review.
-
CD1d-independent activation of mouse and human iNKT cells by bacterial superantigens.Immunol Cell Biol. 2012 Aug;90(7):699-709. doi: 10.1038/icb.2011.90. Epub 2011 Nov 1. Immunol Cell Biol. 2012. PMID: 22041925
-
Turned on by danger: activation of CD1d-restricted invariant natural killer T cells.Immunology. 2012 Sep;137(1):20-7. doi: 10.1111/j.1365-2567.2012.03612.x. Immunology. 2012. PMID: 22734667 Free PMC article. Review.
Cited by
-
Tipping the balance: innate and adaptive immunity in mitochondrial disease.Curr Opin Immunol. 2025 Aug;95:102566. doi: 10.1016/j.coi.2025.102566. Epub 2025 May 26. Curr Opin Immunol. 2025. PMID: 40424975 Review.
-
Gpx4 Regulates Invariant NKT Cell Homeostasis and Function by Preventing Lipid Peroxidation and Ferroptosis.J Immunol. 2024 Oct 1;213(7):941-951. doi: 10.4049/jimmunol.2400246. J Immunol. 2024. PMID: 39158281
-
Metabolic dysregulation impairs lymphocyte function during severe SARS-CoV-2 infection.Commun Biol. 2023 Apr 7;6(1):374. doi: 10.1038/s42003-023-04730-4. Commun Biol. 2023. PMID: 37029220 Free PMC article.
-
METTL14-dependent m6A modification controls iNKT cell development and function.Cell Rep. 2022 Aug 2;40(5):111156. doi: 10.1016/j.celrep.2022.111156. Cell Rep. 2022. PMID: 35926466 Free PMC article.
-
Metabolic regulation of T cell development.Front Immunol. 2022 Jul 25;13:946119. doi: 10.3389/fimmu.2022.946119. eCollection 2022. Front Immunol. 2022. PMID: 35958585 Free PMC article. Review.
References
-
- Blagih J., et al. ., The energy sensor AMPK regulates T cell metabolic adaptation and effector responses in vivo. Immunity 42, 41–54 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
