

The CysLT2R receptor mediates leukotriene C4-driven acute and chronic itch
- PMID: 33753496
- PMCID: PMC8020753
- DOI: 10.1073/pnas.2022087118
The CysLT2R receptor mediates leukotriene C4-driven acute and chronic itch
Abstract
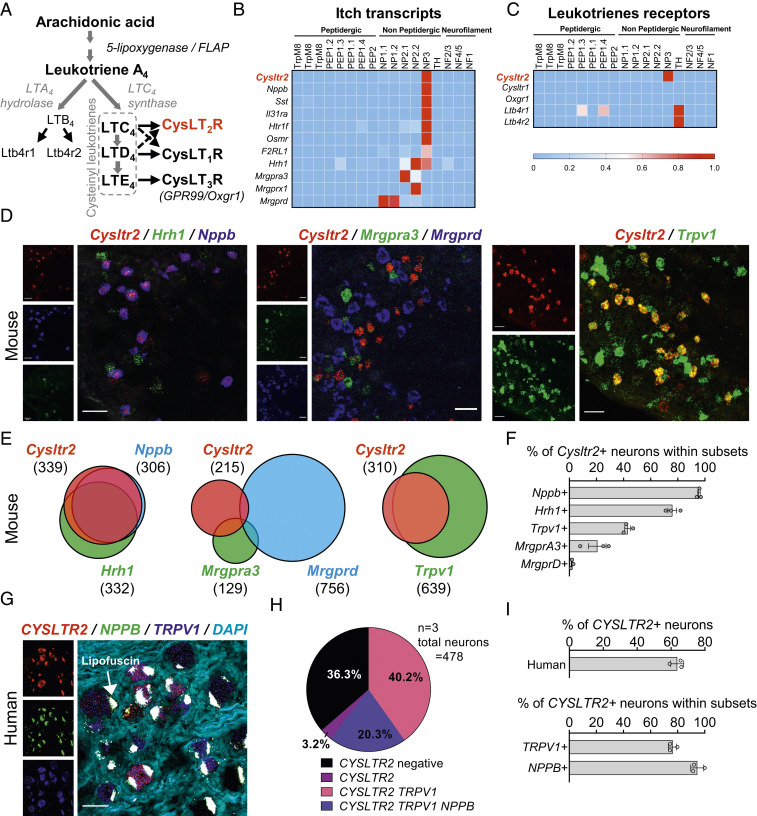
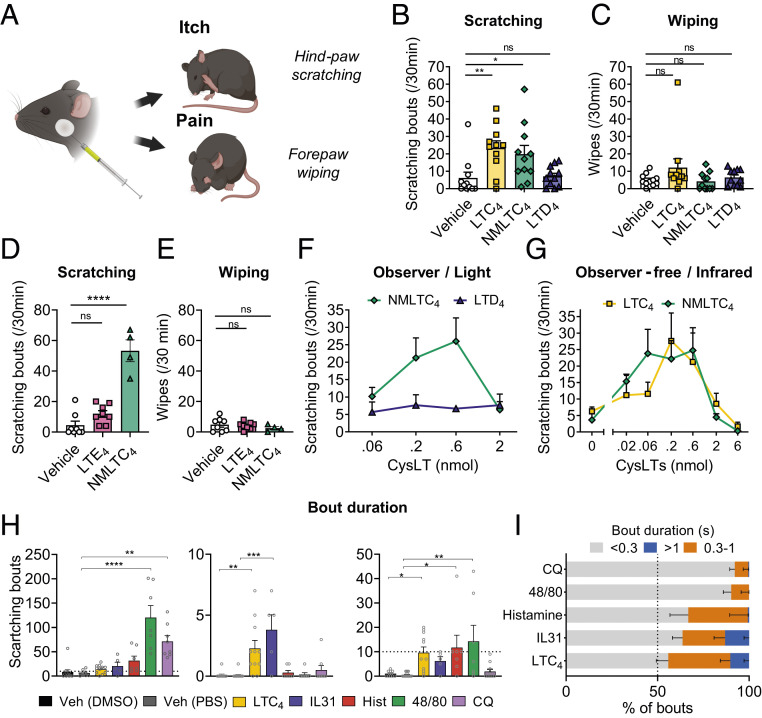
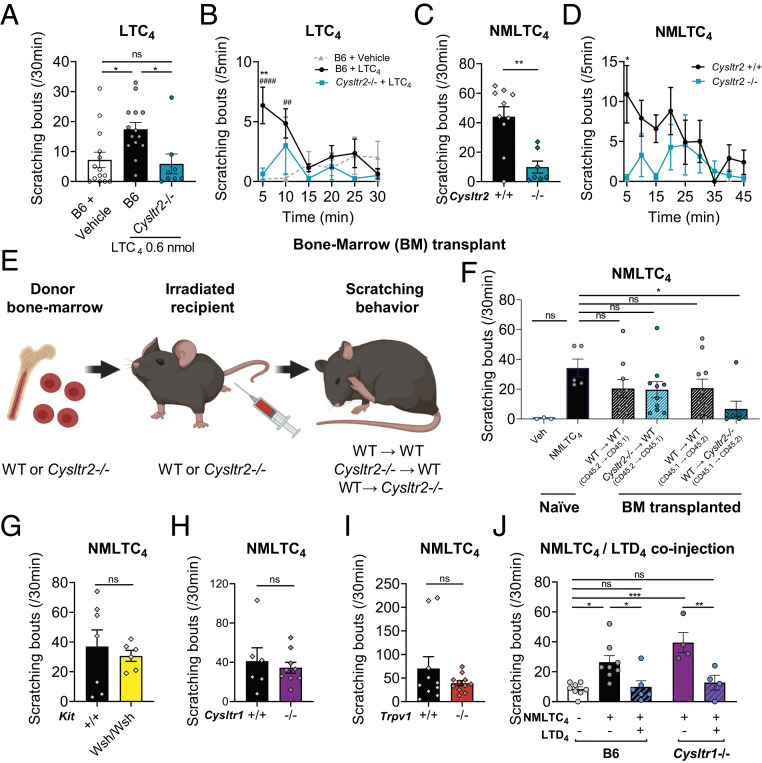
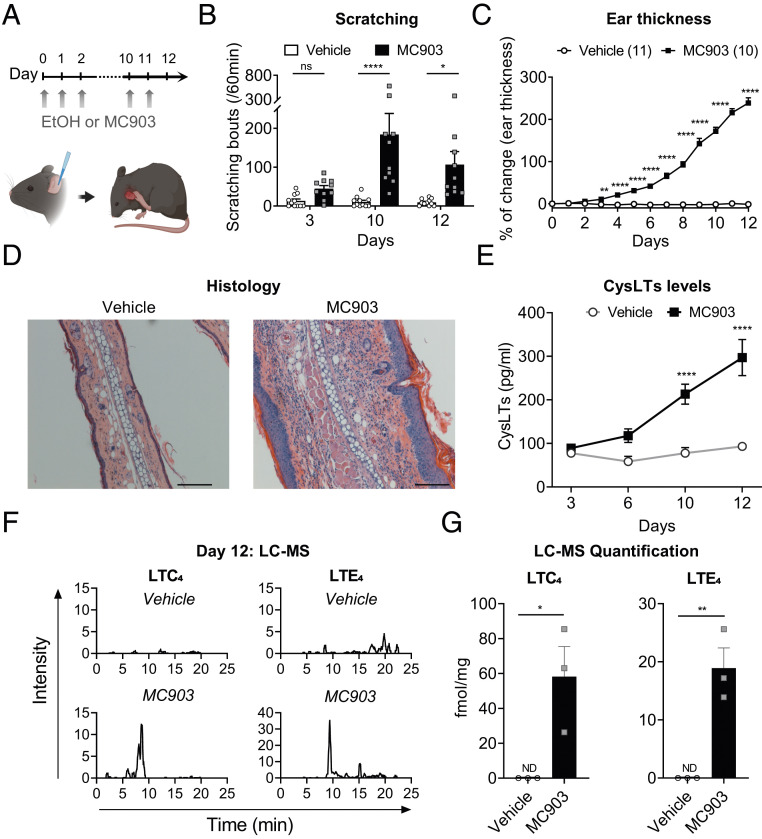
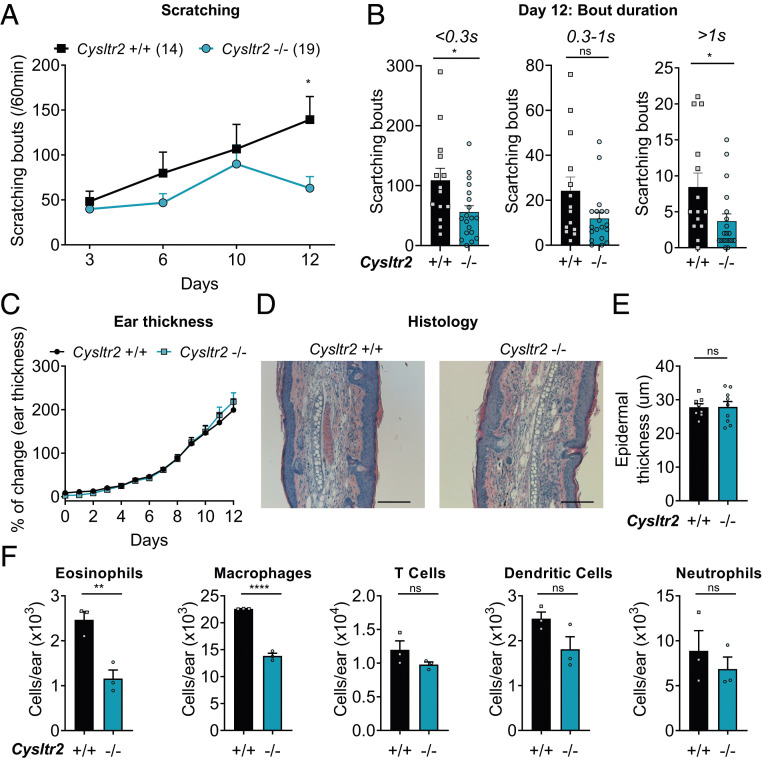
Acute and chronic itch are burdensome manifestations of skin pathologies including allergic skin diseases and atopic dermatitis, but the underlying molecular mechanisms are not well understood. Cysteinyl leukotrienes (CysLTs), comprising LTC4, LTD4, and LTE4, are produced by immune cells during type 2 inflammation. Here, we uncover a role for LTC4 and its signaling through the CysLT receptor 2 (CysLT2R) in itch. Cysltr2 transcript is highly expressed in dorsal root ganglia (DRG) neurons linked to itch in mice. We also detected CYSLTR2 in a broad population of human DRG neurons. Injection of leukotriene C4 (LTC4) or its nonhydrolyzable form NMLTC4, but neither LTD4 nor LTE4, induced dose-dependent itch but not pain behaviors in mice. LTC4-mediated itch differed in bout duration and kinetics from pruritogens histamine, compound 48/80, and chloroquine. NMLTC4-induced itch was abrogated in mice deficient for Cysltr2 or when deficiency was restricted to radioresistant cells. Itch was unaffected in mice deficient for Cysltr1, Trpv1, or mast cells (WSh mice). CysLT2R played a role in itch in the MC903 mouse model of chronic itch and dermatitis, but not in models of dry skin or compound 48/80- or Alternaria-induced itch. In MC903-treated mice, CysLT levels increased in skin over time, and Cysltr2-/- mice showed decreased itch in the chronic phase of inflammation. Collectively, our study reveals that LTC4 acts through CysLT2R as its physiological receptor to induce itch, and CysLT2R contributes to itch in a model of dermatitis. Therefore, targeting CysLT signaling may be a promising approach to treat inflammatory itch.
Keywords: atopic dermatitis; inflammation; itch; neuroimmune; skin.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Kanaoka Y., Austen K. F., Roles of cysteinyl leukotrienes and their receptors in immune cell-related functions. Adv. Immunol. 142, 65–84 (2019). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
