

The number of catalytic cycles in an enzyme's lifetime and why it matters to metabolic engineering
- PMID: 33753504
- PMCID: PMC8020674
- DOI: 10.1073/pnas.2023348118
The number of catalytic cycles in an enzyme's lifetime and why it matters to metabolic engineering
Abstract
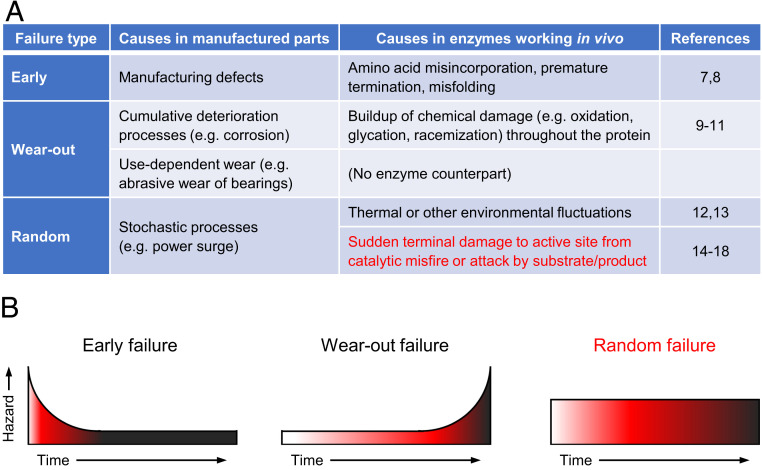
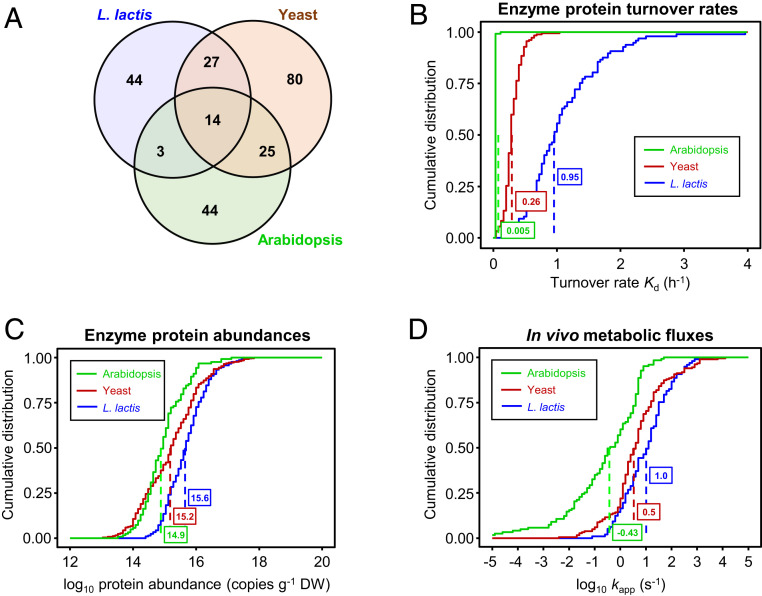
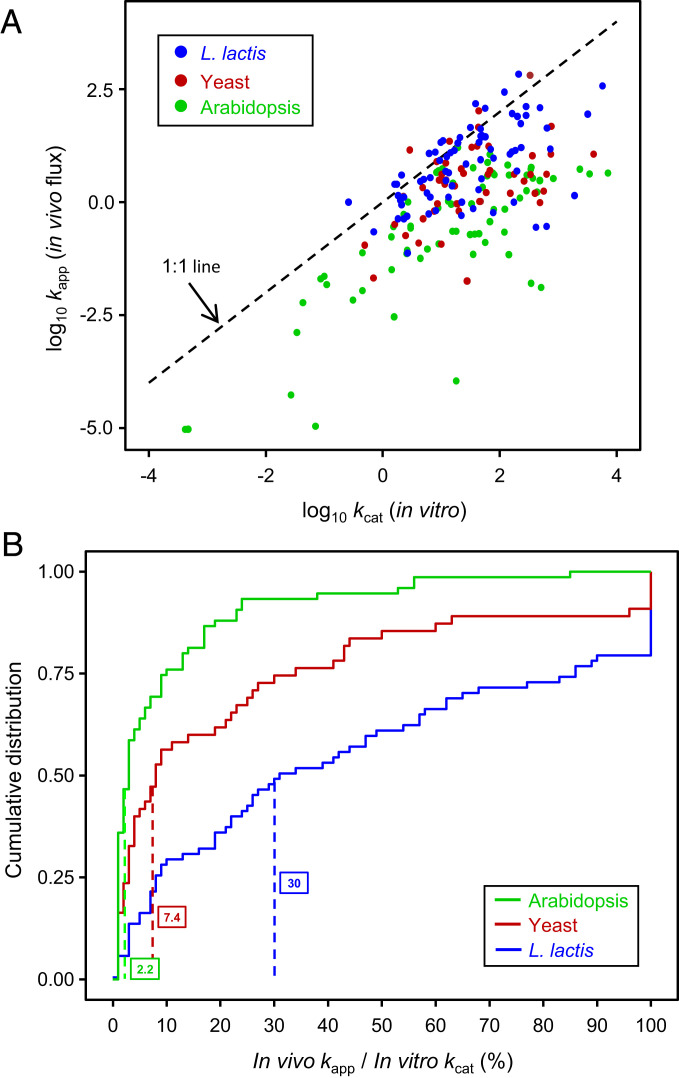
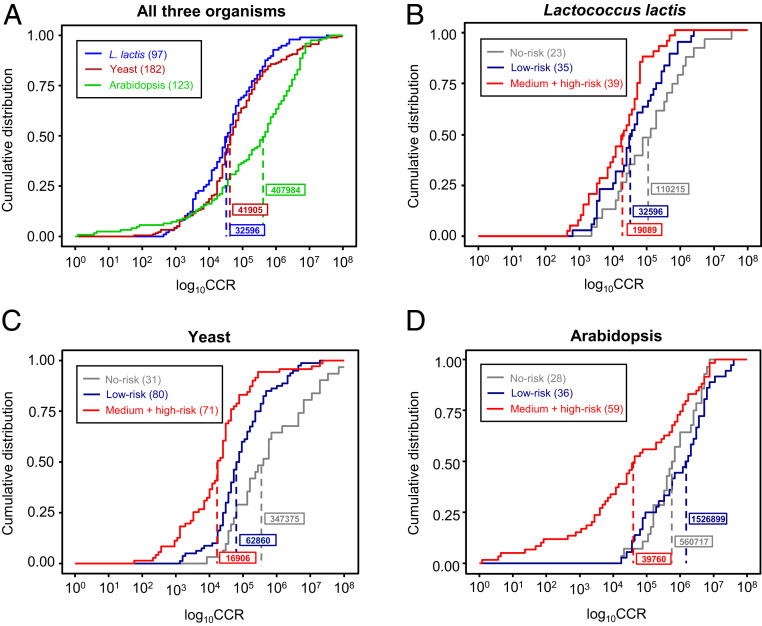
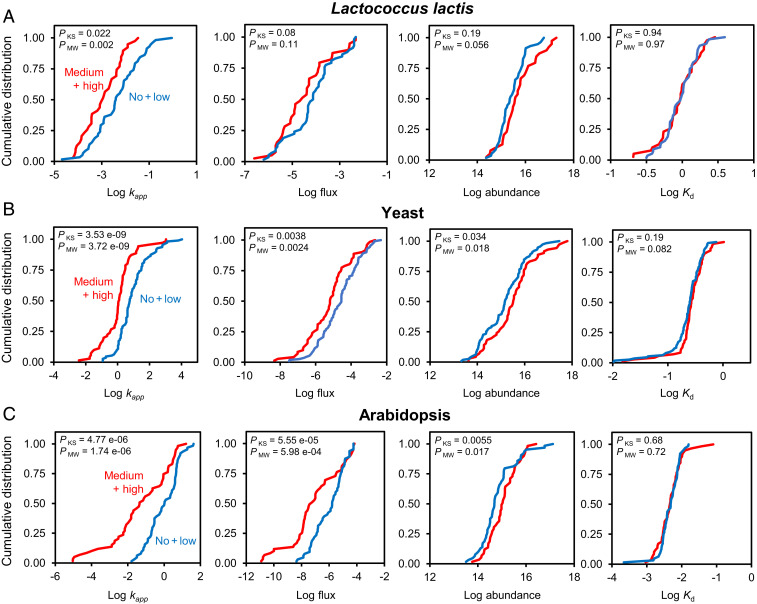
Metabolic engineering uses enzymes as parts to build biosystems for specified tasks. Although a part's working life and failure modes are key engineering performance indicators, this is not yet so in metabolic engineering because it is not known how long enzymes remain functional in vivo or whether cumulative deterioration (wear-out), sudden random failure, or other causes drive replacement. Consequently, enzymes cannot be engineered to extend life and cut the high energy costs of replacement. Guided by catalyst engineering, we adopted catalytic cycles until replacement (CCR) as a metric for enzyme functional life span in vivo. CCR is the number of catalytic cycles that an enzyme mediates in vivo before failure or replacement, i.e., metabolic flux rate/protein turnover rate. We used estimated fluxes and measured protein turnover rates to calculate CCRs for ∼100-200 enzymes each from Lactococcus lactis, yeast, and Arabidopsis CCRs in these organisms had similar ranges (<103 to >107) but different median values (3-4 × 104 in L. lactis and yeast versus 4 × 105 in Arabidopsis). In all organisms, enzymes whose substrates, products, or mechanisms can attack reactive amino acid residues had significantly lower median CCR values than other enzymes. Taken with literature on mechanism-based inactivation, the latter finding supports the proposal that 1) random active-site damage by reaction chemistry is an important cause of enzyme failure, and 2) reactive noncatalytic residues in the active-site region are likely contributors to damage susceptibility. Enzyme engineering to raise CCRs and lower replacement costs may thus be both beneficial and feasible.
Keywords: catalytic cycles; energetic costs; enzyme longevity; protein turnover; synthetic biology.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Way J. C., Collins J. J., Keasling J. D., Silver P. A., Integrating biological redesign: Where synthetic biology came from and where it needs to go. Cell 157, 151–161 (2014). - PubMed
-
- Silver P. A., Way J. C., Arnold F. H., Meyerowitz J. T., Synthetic biology: Engineering explored. Nature 509, 166–167 (2014). - PubMed
-
- Amadi-Echendu J. E., et al., “What is engineering asset management?” in Definitions, Concepts and Scope of Engineering Asset Management, Amadi-Echendu J. E., Brown K., Willett R., Mathew J., Eds. (Springer, 2010), pp. 3–16.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
