

With or Without W? Molecular and Cytogenetic Markers are Not Sufficient for Identification of Environmentally-Induced Sex Reversal in the Bearded Dragon
- PMID: 33756476
- PMCID: PMC8491511
- DOI: 10.1159/000514195
With or Without W? Molecular and Cytogenetic Markers are Not Sufficient for Identification of Environmentally-Induced Sex Reversal in the Bearded Dragon
Erratum in
-
Erratum.Sex Dev. 2021;15(4):282-285. doi: 10.1159/000517966. Epub 2021 Jul 30. Sex Dev. 2021. PMID: 34350896 Free PMC article. No abstract available.
Abstract
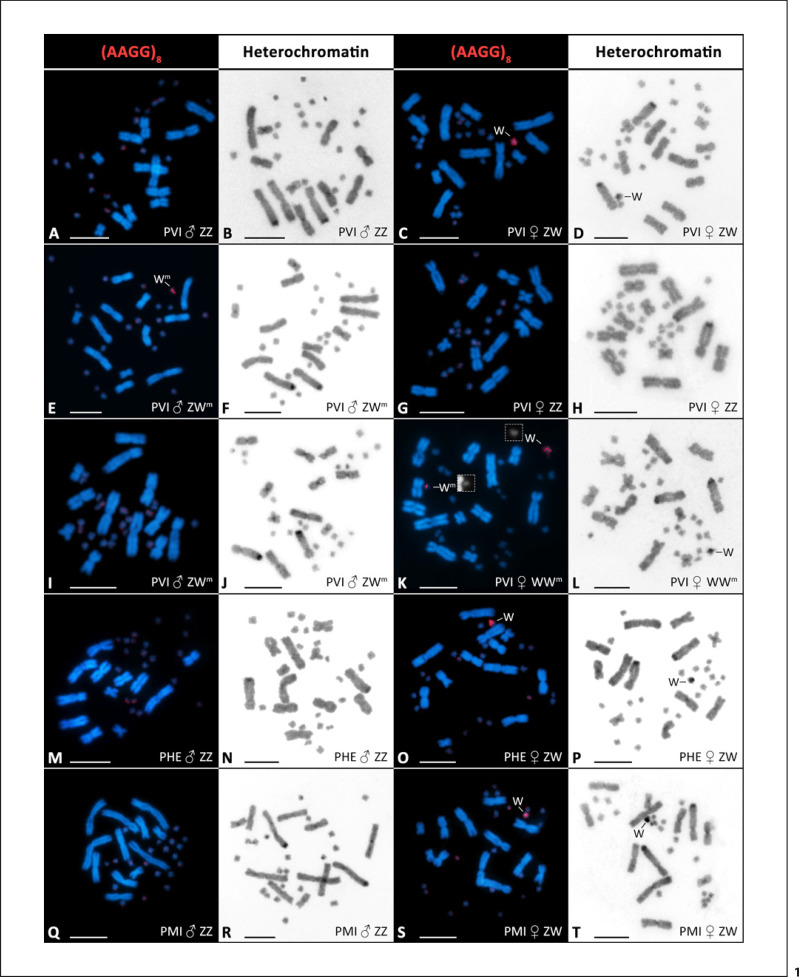
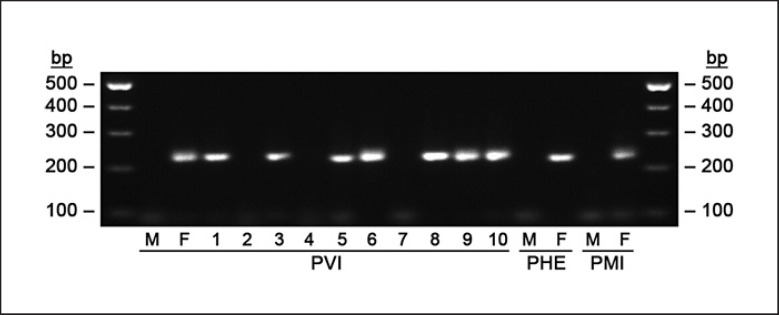
Transitions from environmental sex determination (ESD) to genotypic sex determination (GSD) require an intermediate step of sex reversal, i.e., the production of individuals with a mismatch between the ancestral genotypic and the phenotypic sex. Among amniotes, the sole well-documented transition in this direction was shown in the laboratory in the central bearded dragon, Pogona vitticeps, where very high incubation temperatures led to the production of females with the male-typical (ZZ) genotype. These sex-reversed females then produced offspring whose sex depended on the incubation temperature. Sex-reversed animals identified by molecular and cytogenetic markers were also reported in the field, and their increasing incidence was speculated as a climate warming-driven transition in sex determination. We show that the molecular and cytogenetic markers normally sex-linked in P. vitticeps are also sex-linked in P. henrylawsoni and P. minor, which points to quite ancient sex chromosomes in this lineage. Nevertheless, we demonstrate, based on a crossing experiment with a male bearded dragon who possesses a mismatch between phenotypic sex and genotype, that the used cytogenetic and molecular markers might not be reliable for the identification of sex reversal. Sex reversal should not be considered as the only mechanism causing a mismatch between genetic sex-linked markers and phenotypic sex, which can emerge also by other processes, here most likely by a rare recombination between regions of sex chromosomes which are normally sex-linked. We warn that sex-linked, even apparently for a long evolutionary time, and sex-specific molecular and cytogenetic markers are not a reliable tool for the identification of sex-reversed individuals in a population and that sex reversal has to be verified by other approaches, particularly by observation of the sex ratio of the progeny.
Keywords: Molecular markers; Reversal; Sex chromosomes; Sex linkage; Vertebrates.
© 2021 The Author(s) Published by S. Karger AG, Basel.
Conflict of interest statement
We declare no competing interests in relation to the work.
Figures

Comment in
-
Concerning an Article by Ehl et al.: False Premise Leads to False Conclusions.Sex Dev. 2021;15(4):286-288. doi: 10.1159/000518374. Epub 2021 Jul 29. Sex Dev. 2021. PMID: 34350888 No abstract available.
References
-
- Altmanová M, Rovatsos M, Kratochvíl L, Johnson Pokorná M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae) Biol J Linn Soc. 2016;118((3)):618–33.
-
- Altmanová M, Rovatsos M, Johnson Pokorná M, Veselý M, Wagner F, Kratochvíl L. All iguana families with the exception of basilisks share sex chromosomes. Zoology. 2018;126:98–102. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215((3)):403–10. - PubMed
-
- Castelli MA, Georges A, Cherryh C, Rosauer DF, Sarre SS, Contador‐Kelsall I, et al. Evolving thermal thresholds explain the distribution of temperature sex reversal in an Australian dragon lizard. Divers Distrib. 2020 ..in press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
