

Identification of deleterious recessive haplotypes and candidate deleterious recessive mutations in Japanese Black cattle
- PMID: 33758295
- PMCID: PMC7988166
- DOI: 10.1038/s41598-021-86225-y
Identification of deleterious recessive haplotypes and candidate deleterious recessive mutations in Japanese Black cattle
Abstract
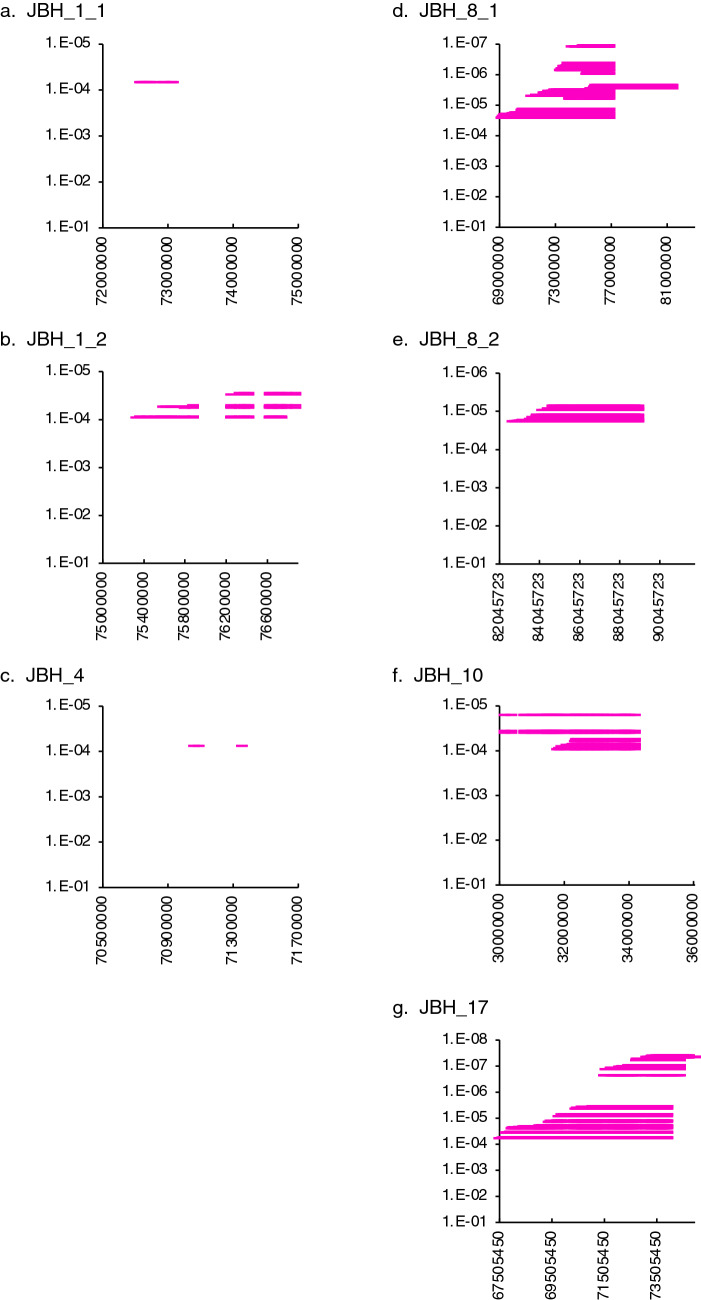
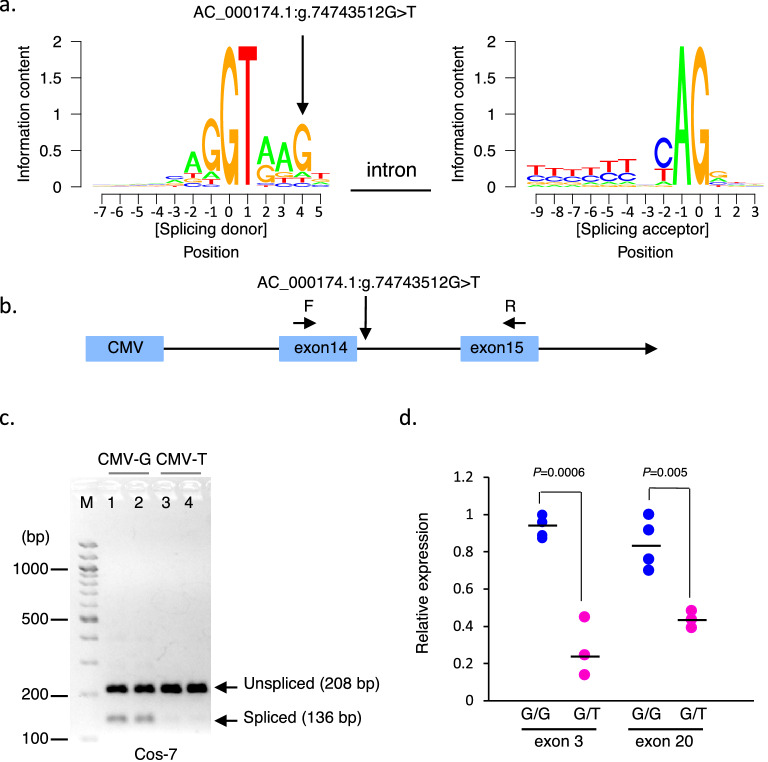
Intensive use of a few elite sires has increased the risk of the manifestation of deleterious recessive traits in cattle. Substantial genotyping data gathered using single-nucleotide polymorphism (SNP) arrays have identified the haplotypes with homozygous deficiency, which may compromise survival. We developed Japanese Black cattle haplotypes (JBHs) using SNP array data (4843 individuals) and identified deleterious recessive haplotypes using exome sequencing of 517 sires. We identified seven JBHs with homozygous deficiency. JBH_10 and JBH_17 were associated with the resuming of estrus after artificial insemination, indicating that these haplotypes carried deleterious mutations affecting embryonic survival. The exome data of 517 Japanese Black sires revealed that AC_000165.1:g.85341291C>G of IARS in JBH_8_2, AC_000174.1:g.74743512G>T of CDC45 in JBH_17, and a copy variation region (CNVR_27) of CLDN16 in JBH_1_1 and JBH_1_2 were the candidate mutations. A novel variant AC_000174.1:g.74743512G>T of CDC45 in JBH_17 was located in a splicing donor site at a distance of 5 bp, affecting pre-mRNA splicing. Mating between heterozygotes of JBH_17 indicated that homozygotes carrying the risk allele died around the blastocyst stage. Analysis of frequency of the CDC45 risk allele revealed that its carriers were widespread throughout the tested Japanese Black cattle population. Our approach can effectively manage the inheritance of recessive risk alleles in a breeding population.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Namikawa, K. Japanese beef cattle: Historical breeding processes of japanese beef cattle and preservation of genetic resources as economic farm animal (in Japanese): Wagyu Registry Association. Wagyu (1992).
-
- Ogawa H. Factor XIII deficiency in Japanese Black cattle (in Japanese) J. Vet. Med. 1996;49:411–412.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
