

Microglial identity and inflammatory responses are controlled by the combined effects of neurons and astrocytes
- PMID: 33761343
- PMCID: PMC7994374
- DOI: 10.1016/j.celrep.2021.108882
Microglial identity and inflammatory responses are controlled by the combined effects of neurons and astrocytes
Abstract
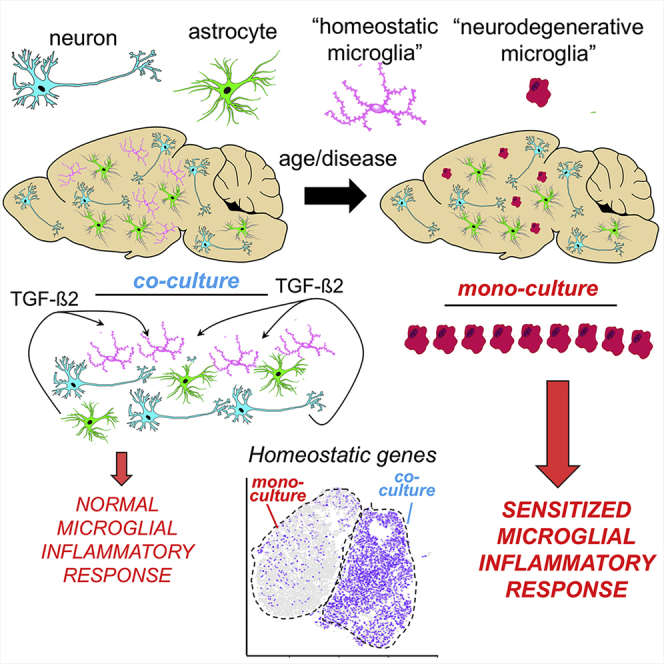
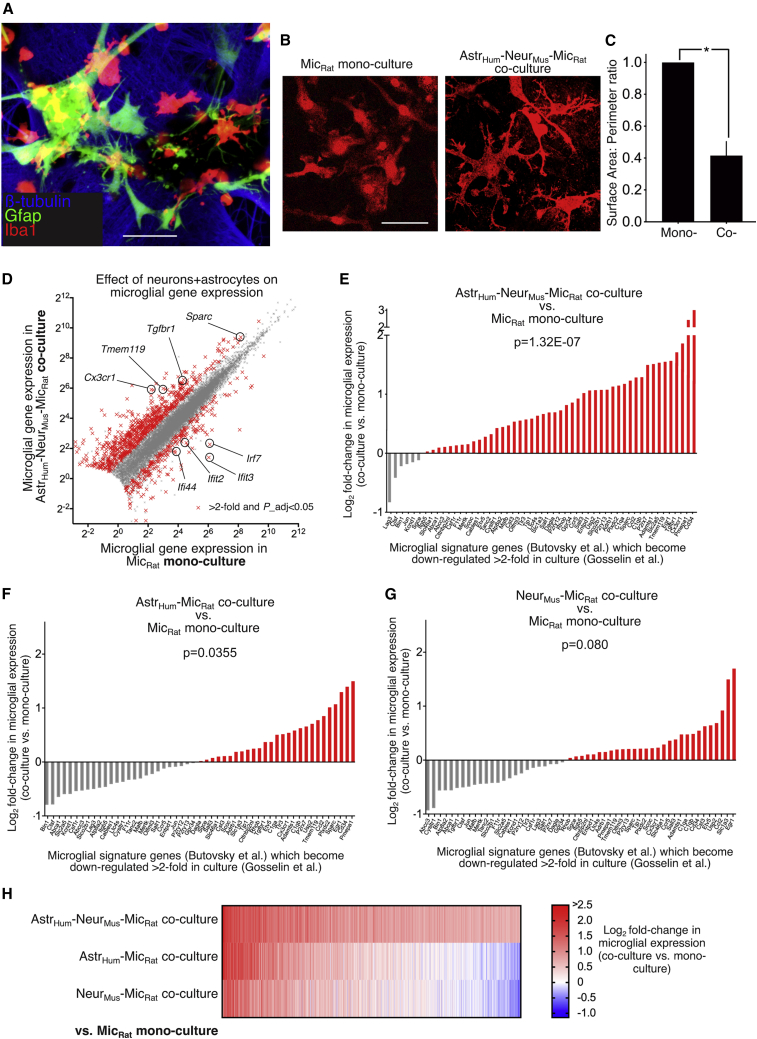
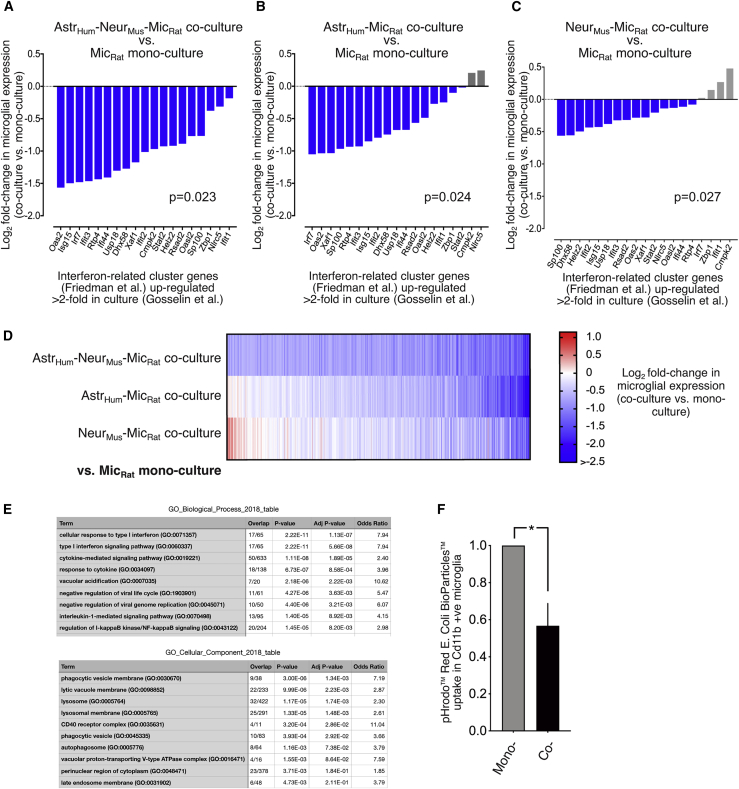
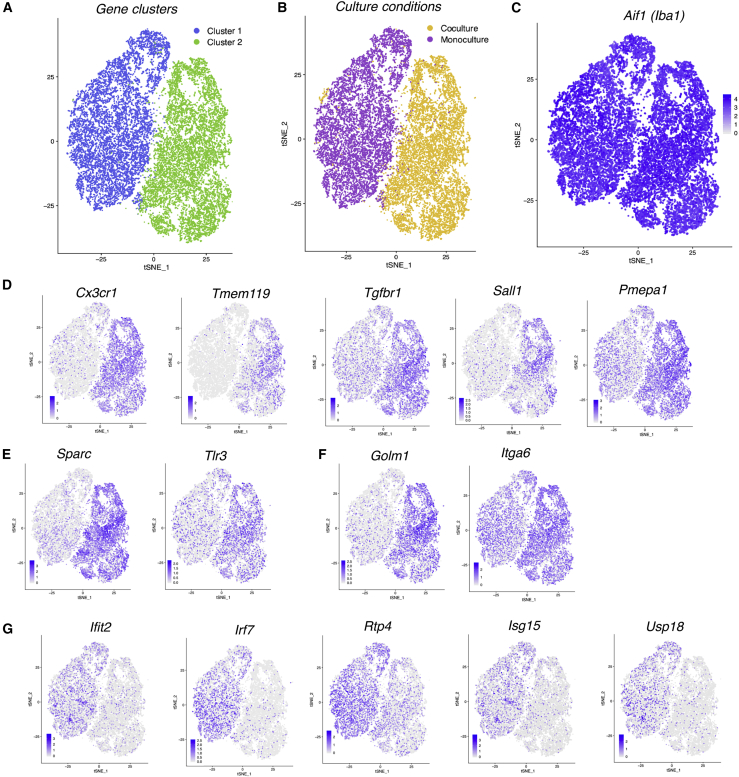
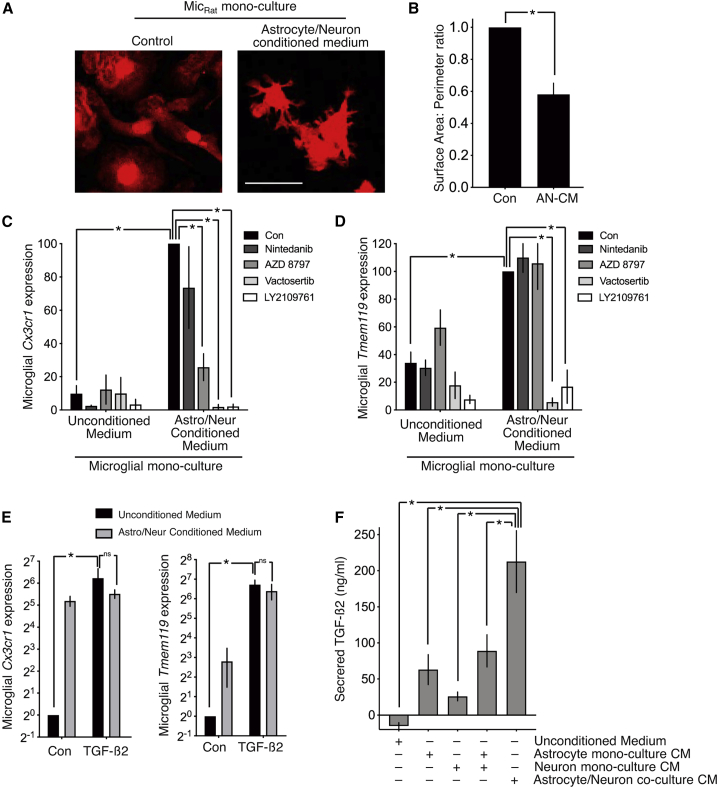
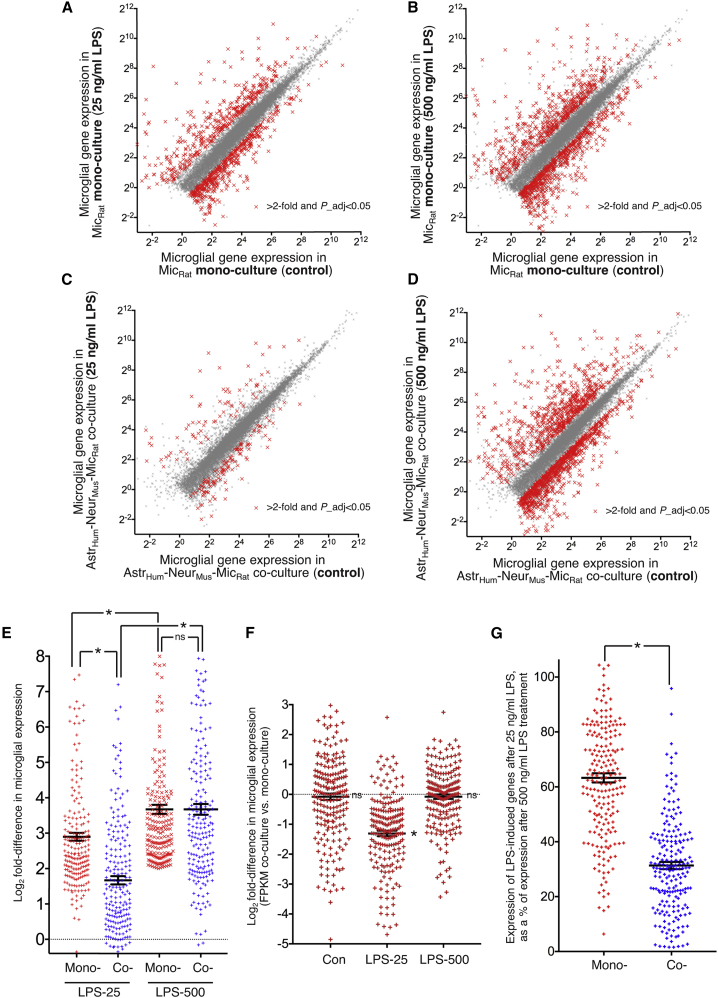
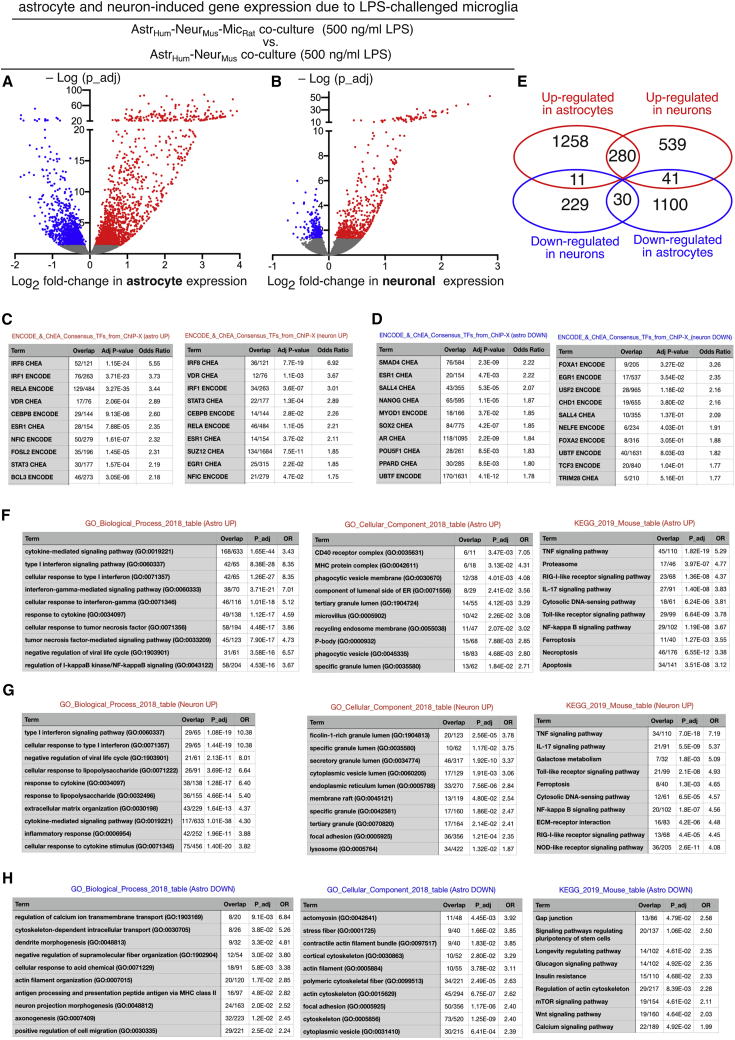
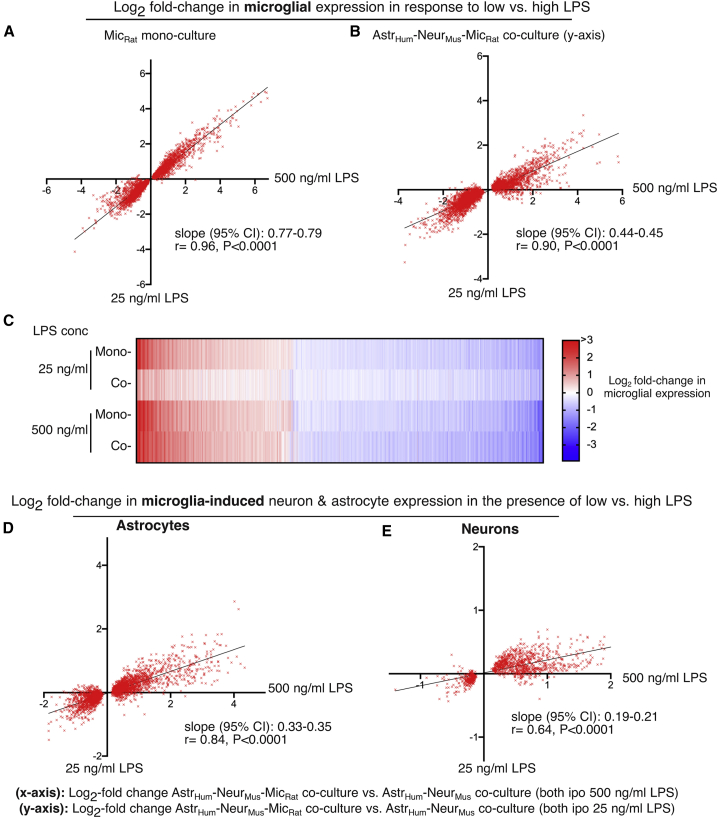
Microglia, brain-resident macrophages, require instruction from the CNS microenvironment to maintain their identity and morphology and regulate inflammatory responses, although what mediates this is unclear. Here, we show that neurons and astrocytes cooperate to promote microglial ramification, induce expression of microglial signature genes ordinarily lost in vitro and in age and disease in vivo, and repress infection- and injury-associated gene sets. The influence of neurons and astrocytes separately on microglia is weak, indicative of synergies between these cell types, which exert their effects via a mechanism involving transforming growth factor β2 (TGF-β2) signaling. Neurons and astrocytes also combine to provide immunomodulatory cues, repressing primed microglial responses to weak inflammatory stimuli (without affecting maximal responses) and consequently limiting the feedback effects of inflammation on the neurons and astrocytes themselves. These findings explain why microglia isolated ex vivo undergo de-differentiation and inflammatory deregulation and point to how disease- and age-associated changes may be counteracted.
Keywords: RNA-seq; ageing; astrocytes; microglia; neurodegeneration; neurons; signal transduction; transcriptomics.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Atg7 deficiency in microglia drives an altered transcriptomic profile associated with an impaired neuroinflammatory response.Mol Brain. 2021 Jun 3;14(1):87. doi: 10.1186/s13041-021-00794-7. Mol Brain. 2021. PMID: 34082793 Free PMC article.
-
Astrocyte-derived CCL2 participates in surgery-induced cognitive dysfunction and neuroinflammation via evoking microglia activation.Behav Brain Res. 2017 Aug 14;332:145-153. doi: 10.1016/j.bbr.2017.05.066. Epub 2017 Jun 3. Behav Brain Res. 2017. PMID: 28587818
-
The neuron-astrocyte-microglia triad in normal brain ageing and in a model of neuroinflammation in the rat hippocampus.PLoS One. 2012;7(9):e45250. doi: 10.1371/journal.pone.0045250. Epub 2012 Sep 18. PLoS One. 2012. PMID: 23028880 Free PMC article.
-
Activation of microglia and astrocytes: a roadway to neuroinflammation and Alzheimer's disease.Inflammopharmacology. 2019 Aug;27(4):663-677. doi: 10.1007/s10787-019-00580-x. Epub 2019 Mar 14. Inflammopharmacology. 2019. PMID: 30874945 Review.
-
Hypothalamic Microglial Heterogeneity and Signature under High Fat Diet-Induced Inflammation.Int J Mol Sci. 2021 Feb 24;22(5):2256. doi: 10.3390/ijms22052256. Int J Mol Sci. 2021. PMID: 33668314 Free PMC article. Review.
Cited by
-
Plasma derived extracellular vesicle biomarkers of microglia activation in an experimental stroke model.J Neuroinflammation. 2023 Jan 31;20(1):20. doi: 10.1186/s12974-023-02708-x. J Neuroinflammation. 2023. PMID: 36721258 Free PMC article.
-
Agomir-331 Suppresses Reactive Gliosis and Neuroinflammation after Traumatic Brain Injury.Cells. 2023 Oct 11;12(20):2429. doi: 10.3390/cells12202429. Cells. 2023. PMID: 37887272 Free PMC article.
-
The Meninges as CNS Interfaces and the Roles of Meningeal Macrophages.Biomolecules. 2025 Mar 28;15(4):497. doi: 10.3390/biom15040497. Biomolecules. 2025. PMID: 40305192 Free PMC article. Review.
-
Functional genomics in stem cell models: considerations and applications.Front Cell Dev Biol. 2023 Jul 24;11:1236553. doi: 10.3389/fcell.2023.1236553. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37554308 Free PMC article. Review.
-
IRF3 regulates neuroinflammatory responses and the expression of genes associated with Alzheimer's disease.bioRxiv [Preprint]. 2024 Mar 12:2024.03.08.582968. doi: 10.1101/2024.03.08.582968. bioRxiv. 2024. Update in: J Neuroinflammation. 2024 Aug 30;21(1):212. doi: 10.1186/s12974-024-03203-7. PMID: 38654824 Free PMC article. Updated. Preprint.
References
-
- Baxter P.S., Martel M.A., McMahon A., Kind P.C., Hardingham G.E. Pituitary adenylate cyclase-activating peptide induces long-lasting neuroprotection through the induction of activity-dependent signaling via the cyclic AMP response element-binding protein-regulated transcription co-activator 1. J. Neurochem. 2011;118:365–378. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
