

Human MC4R variants affect endocytosis, trafficking and dimerization revealing multiple cellular mechanisms involved in weight regulation
- PMID: 33761344
- PMCID: PMC7994375
- DOI: 10.1016/j.celrep.2021.108862
Human MC4R variants affect endocytosis, trafficking and dimerization revealing multiple cellular mechanisms involved in weight regulation
Abstract
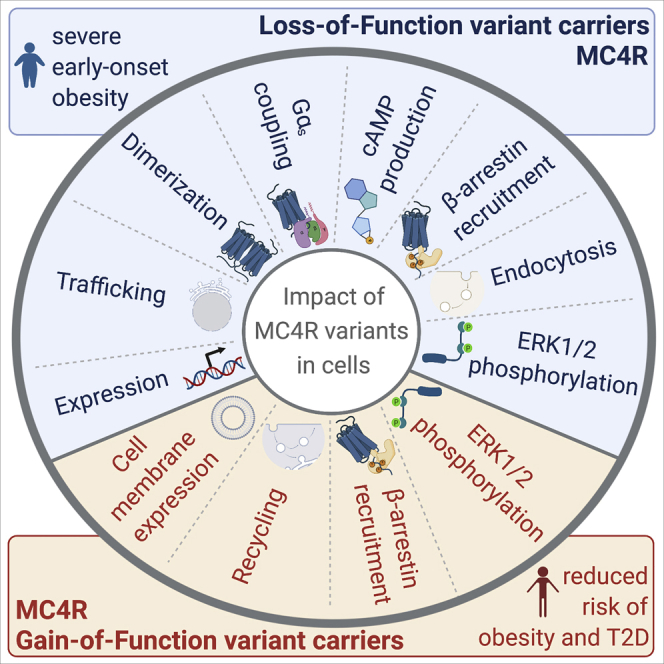
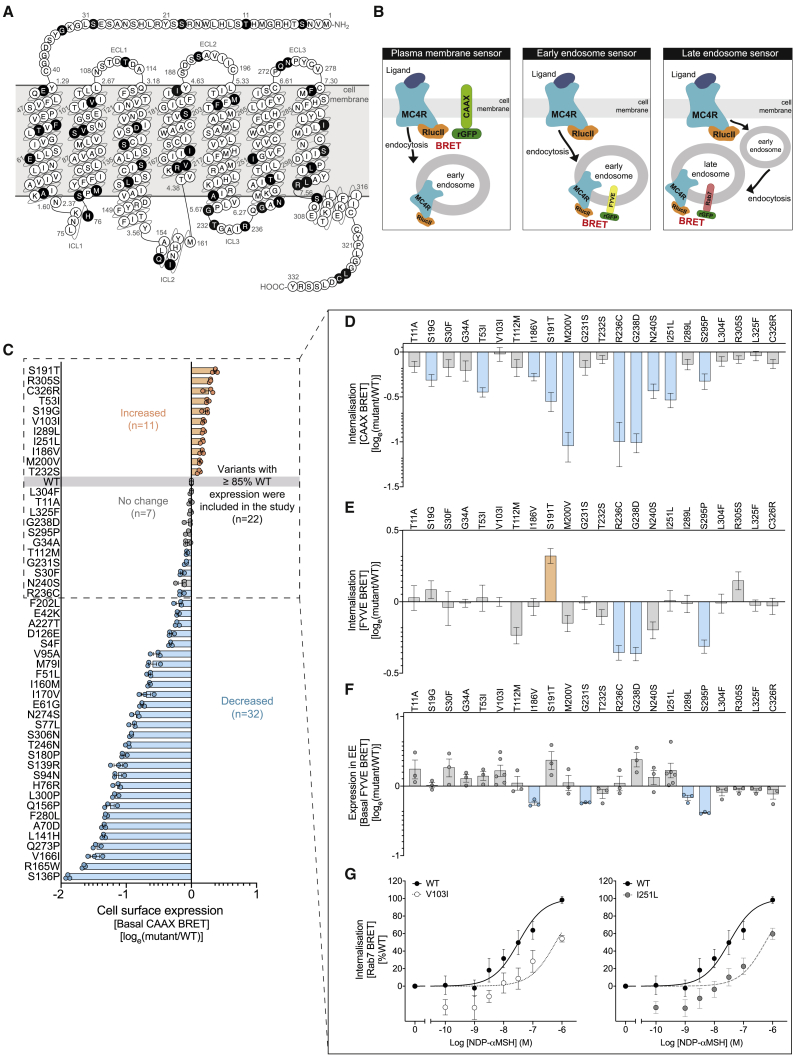
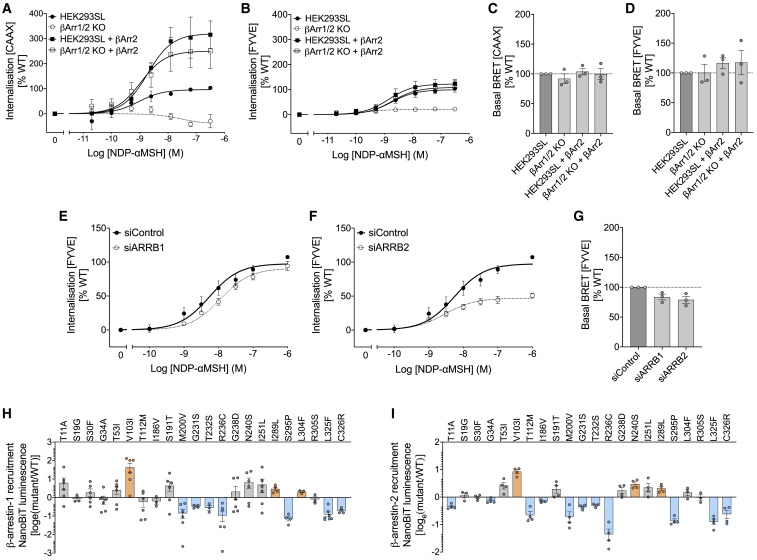
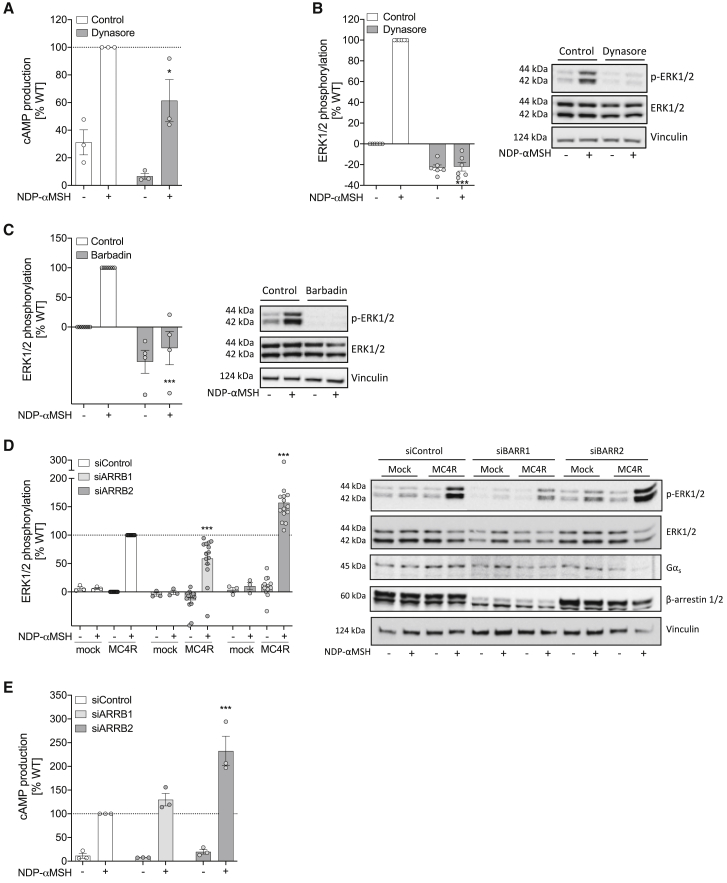
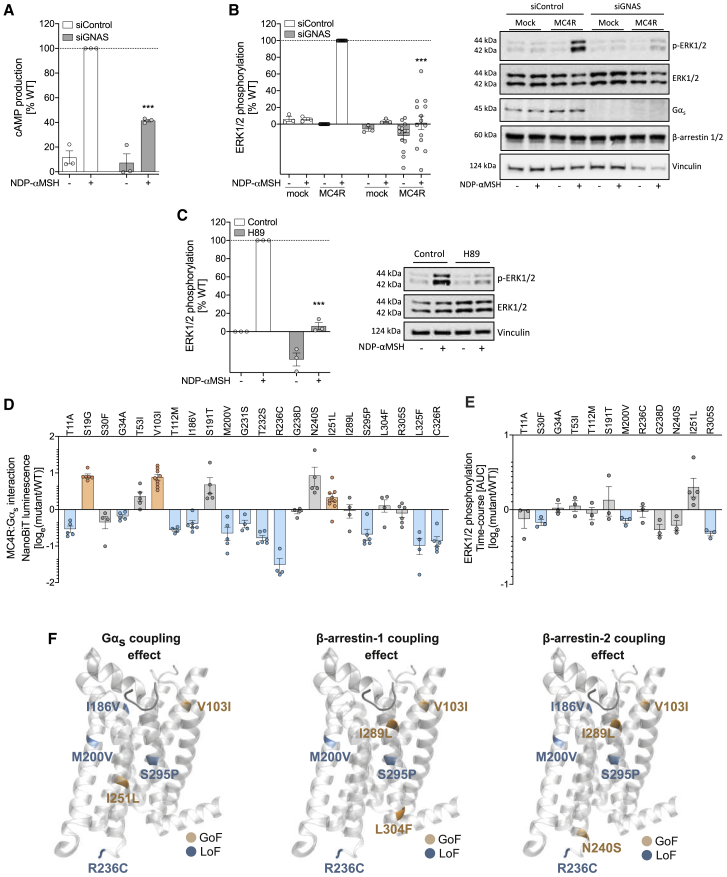
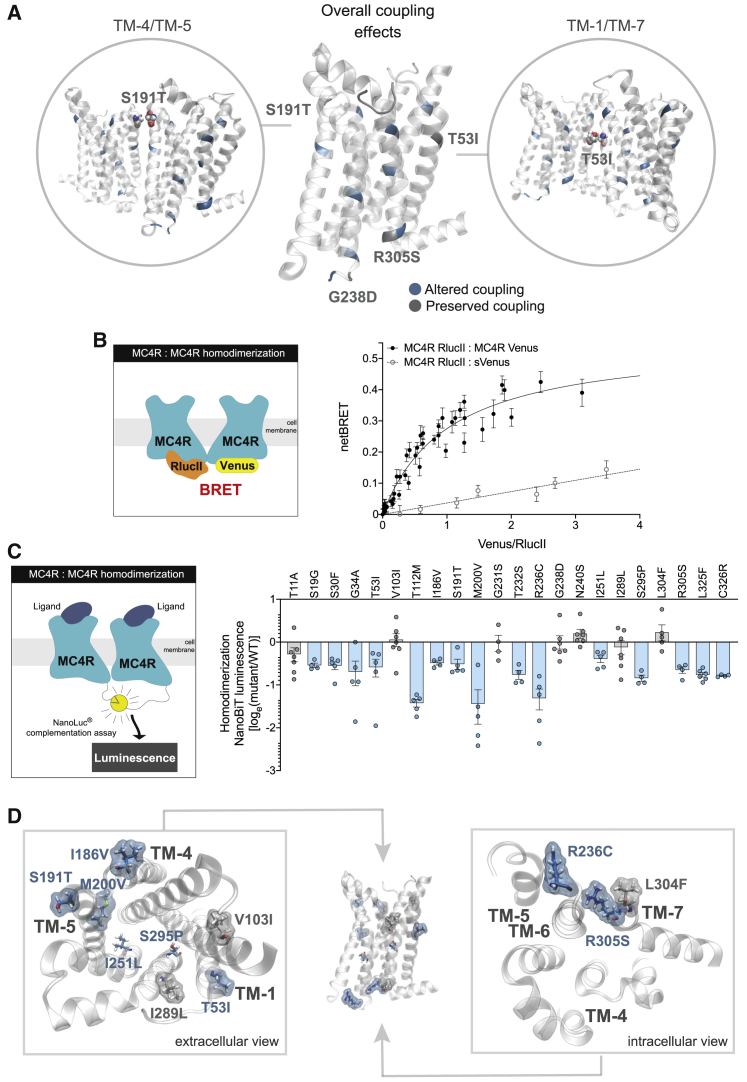
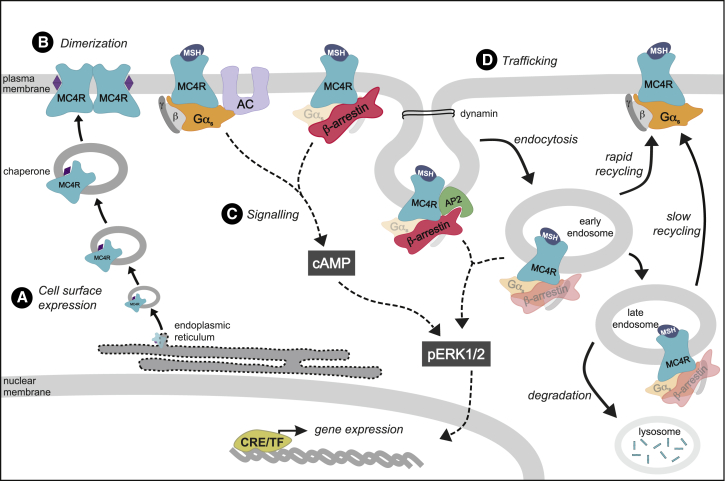
The Melanocortin-4 Receptor (MC4R) plays a pivotal role in energy homeostasis. We used human MC4R mutations associated with an increased or decreased risk of obesity to dissect mechanisms that regulate MC4R function. Most obesity-associated mutations impair trafficking to the plasma membrane (PM), whereas obesity-protecting mutations either accelerate recycling to the PM or decrease internalization, resulting in enhanced signaling. MC4R mutations that do not affect canonical Gαs protein-mediated signaling, previously considered to be non-pathogenic, nonetheless disrupt agonist-induced internalization, β-arrestin recruitment, and/or coupling to Gαs, establishing their causal role in severe obesity. Structural mapping reveals ligand-accessible sites by which MC4R couples to effectors and residues involved in the homodimerization of MC4R, which is disrupted by multiple obesity-associated mutations. Human genetic studies reveal that endocytosis, intracellular trafficking, and homodimerization regulate MC4R function to a level that is physiologically relevant, supporting the development of chaperones, agonists, and allosteric modulators of MC4R for weight loss therapy.
Keywords: GPCRs; Gα(s); MC4R; MSH; melanocortin; obesity; therapy; weight loss; β-arrestin.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Regulation of melanocortin-4 receptor signaling: agonist-mediated desensitization and internalization.Endocrinology. 2003 Apr;144(4):1301-14. doi: 10.1210/en.2002-220931. Endocrinology. 2003. PMID: 12639913
-
Human Gain-of-Function MC4R Variants Show Signaling Bias and Protect against Obesity.Cell. 2019 Apr 18;177(3):597-607.e9. doi: 10.1016/j.cell.2019.03.044. Cell. 2019. PMID: 31002796 Free PMC article.
-
MC4R Variants Modulate α-MSH and Setmelanotide Induced Cellular Signaling at Multiple Levels.J Clin Endocrinol Metab. 2024 Sep 16;109(10):2452-2466. doi: 10.1210/clinem/dgae210. J Clin Endocrinol Metab. 2024. PMID: 38567654 Free PMC article.
-
Constitutive activity in melanocortin-4 receptor: biased signaling of inverse agonists.Adv Pharmacol. 2014;70:135-54. doi: 10.1016/B978-0-12-417197-8.00005-5. Adv Pharmacol. 2014. PMID: 24931195 Review.
-
Structural Complexity and Plasticity of Signaling Regulation at the Melanocortin-4 Receptor.Int J Mol Sci. 2020 Aug 10;21(16):5728. doi: 10.3390/ijms21165728. Int J Mol Sci. 2020. PMID: 32785054 Free PMC article. Review.
Cited by
-
A human obesity-associated MC4R mutation with defective Gq/11α signaling leads to hyperphagia in mice.J Clin Invest. 2024 Jan 4;134(4):e165418. doi: 10.1172/JCI165418. J Clin Invest. 2024. PMID: 38175730 Free PMC article.
-
Current Treatments for Patients with Genetic Obesity.J Clin Res Pediatr Endocrinol. 2023 May 29;15(2):108-119. doi: 10.4274/jcrpe.galenos.2023.2023-3-2. Epub 2023 May 16. J Clin Res Pediatr Endocrinol. 2023. PMID: 37191347 Free PMC article. Review.
-
Detecting genetic epistasis by differential departure from independence.Mol Genet Genomics. 2022 Jul;297(4):911-924. doi: 10.1007/s00438-022-01893-3. Epub 2022 May 23. Mol Genet Genomics. 2022. PMID: 35606612
-
Unexpected identification of obesity-associated mutations in LEP and MC4R genes in patients with anorexia nervosa.Sci Rep. 2024 Mar 25;14(1):7067. doi: 10.1038/s41598-024-57517-w. Sci Rep. 2024. PMID: 38528040 Free PMC article.
-
Identification and molecular characterization of missense mutations in orphan G protein-coupled receptor GPR61 occurring in severe obesity.Mol Pharmacol. 2025 Apr;107(4):100026. doi: 10.1016/j.molpha.2025.100026. Epub 2025 Mar 4. Mol Pharmacol. 2025. PMID: 40133016 Free PMC article.
References
-
- Biebermann H., Krude H., Elsner A., Chubanov V., Gudermann T., Grüters A. Autosomal-dominant mode of inheritance of a melanocortin-4 receptor mutation in a patient with severe early-onset obesity is due to a dominant-negative effect caused by receptor dimerization. Diabetes. 2003;52:2984–2988. - PubMed
-
- Capra V., Mauri M., Guzzi F., Busnelli M., Accomazzo M.R., Gaussem P., Nisar S.P., Mundell S.J., Parenti M., Rovati G.E. Impaired thromboxane receptor dimerization reduces signaling efficiency: A potential mechanism for reduced platelet function in vivo. Biochem. Pharmacol. 2017;124:43–56. - PubMed
-
- Clément K., Biebermann H., Farooqi I.S., Van der Ploeg L., Wolters B., Poitou C., Puder L., Fiedorek F., Gottesdiener K., Kleinau G. MC4R agonism promotes durable weight loss in patients with leptin receptor deficiency. Nat. Med. 2018;24:551–555. - PubMed
-
- Clément K., van den Akker E., Argente J., Bahm A., Chung W.K., Connors H., De Waele K., Farooqi I.S., Gonneau-Lejeune J., Gordon G. Efficacy and safety of setmelanotide, an MC4R agonist, in individuals with severe obesity due to LEPR or POMC deficiency: single-arm, open-label, multicentre, phase 3 trials. Lancet Diabetes Endocrinol. 2020;8:960–970. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
