

Ca2+ signals critical for egress and gametogenesis in malaria parasites depend on a multipass membrane protein that interacts with PKG
- PMID: 33762339
- PMCID: PMC7990342
- DOI: 10.1126/sciadv.abe5396
Ca2+ signals critical for egress and gametogenesis in malaria parasites depend on a multipass membrane protein that interacts with PKG
Abstract
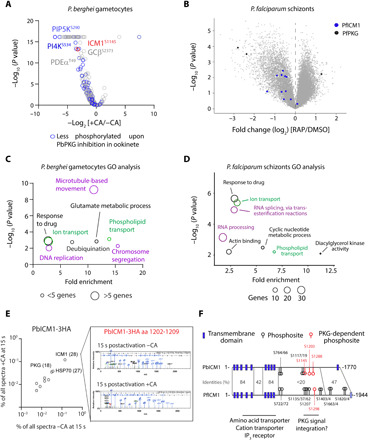
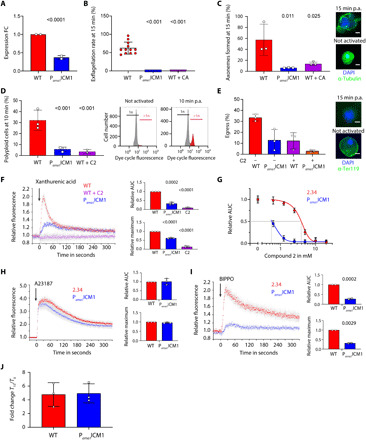
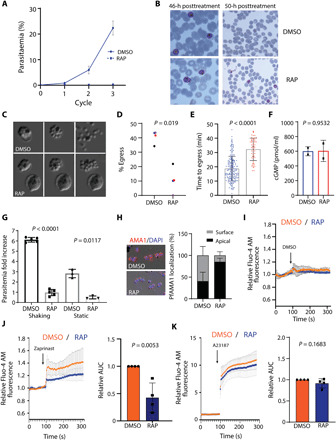
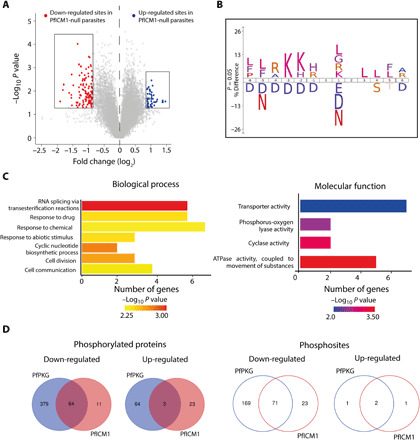
Calcium signaling regulated by the cGMP-dependent protein kinase (PKG) controls key life cycle transitions in the malaria parasite. However, how calcium is mobilized from intracellular stores in the absence of canonical calcium channels in Plasmodium is unknown. Here, we identify a multipass membrane protein, ICM1, with homology to transporters and calcium channels that is tightly associated with PKG in both asexual blood stages and transmission stages. Phosphoproteomic analyses reveal multiple ICM1 phosphorylation events dependent on PKG activity. Stage-specific depletion of Plasmodium berghei ICM1 prevents gametogenesis due to a block in intracellular calcium mobilization, while conditional loss of Plasmodium falciparum ICM1 is detrimental for the parasite resulting in severely reduced calcium mobilization, defective egress, and lack of invasion. Our findings suggest that ICM1 is a key missing link in transducing PKG-dependent signals and provide previously unknown insights into atypical calcium homeostasis in malaria parasites essential for pathology and disease transmission.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures

References
-
- WHO, World malaria report (2019).
-
- Brochet M., Billker O., Calcium signalling in malaria parasites. Mol. Microbiol. 100, 397–408 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
