

Phenotypic diversity and metabolic specialization of renal endothelial cells
- PMID: 33767431
- PMCID: PMC7993417
- DOI: 10.1038/s41581-021-00411-9
Phenotypic diversity and metabolic specialization of renal endothelial cells
Abstract
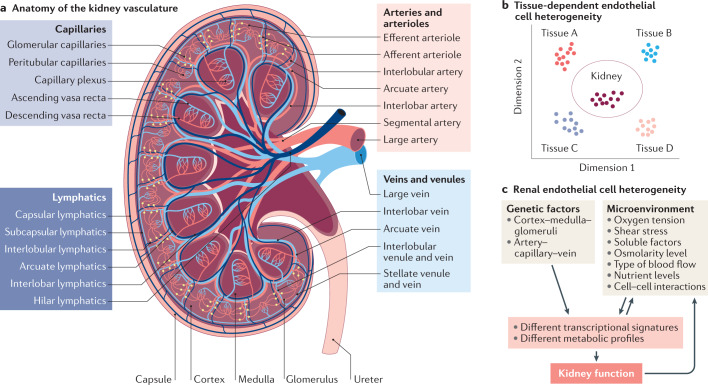
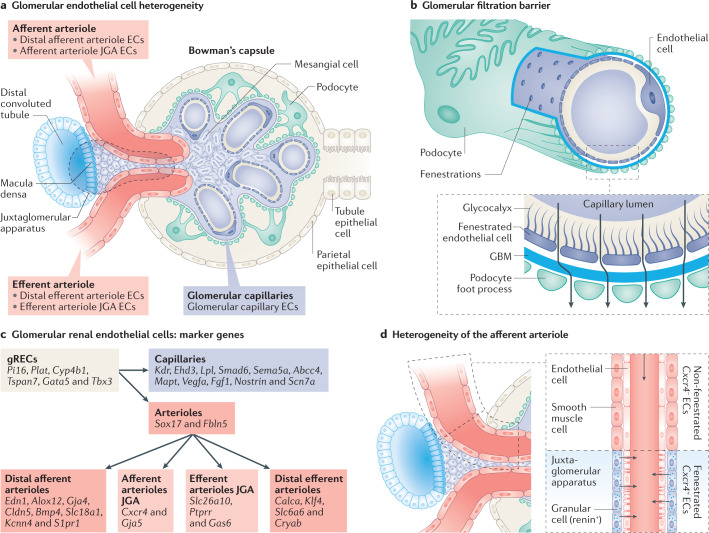
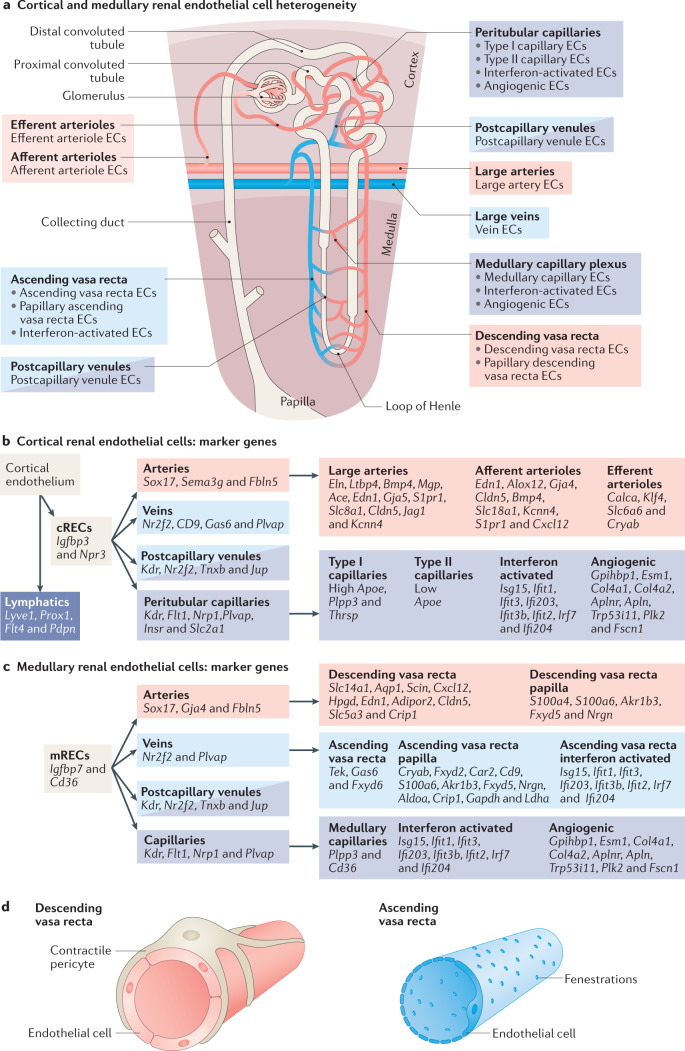
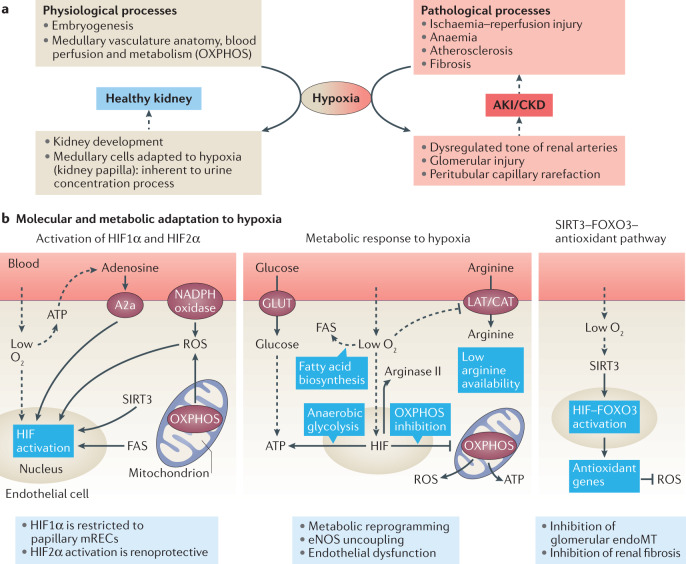
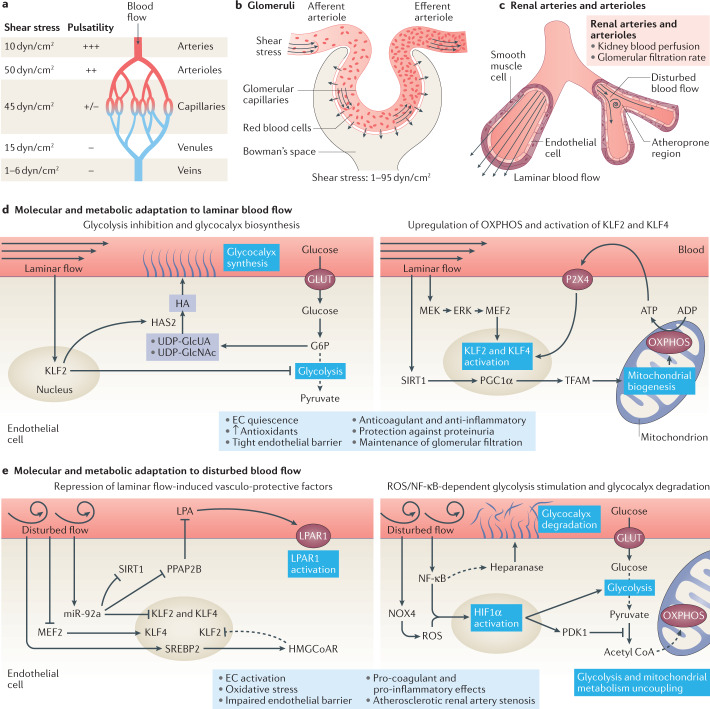
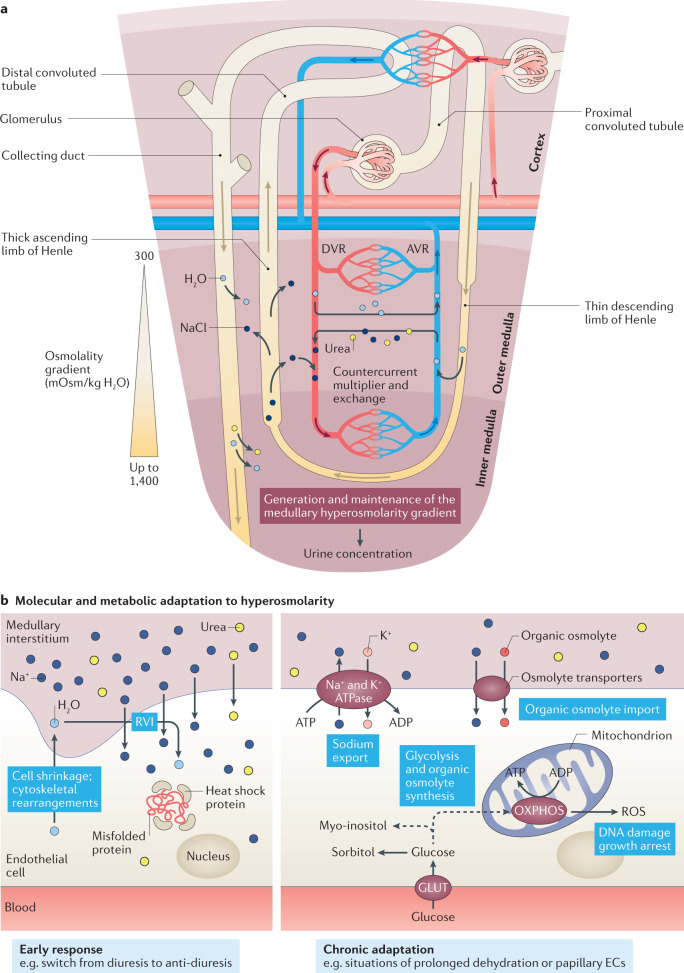
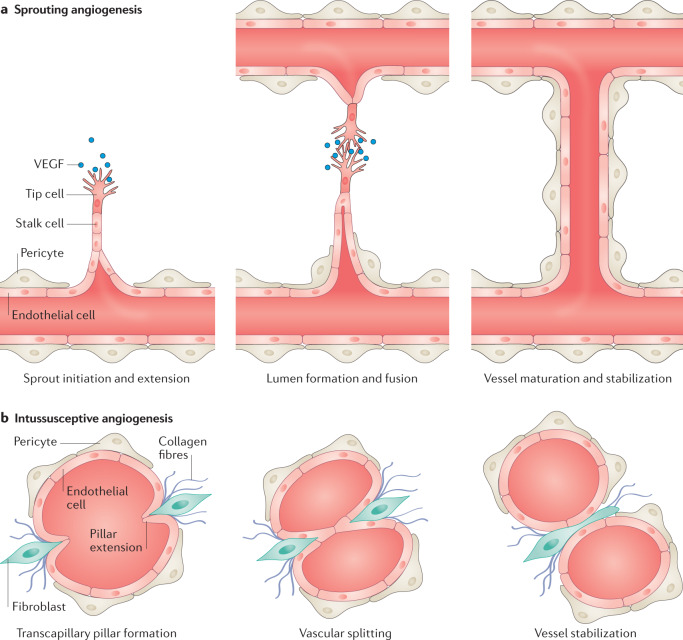
Complex multicellular life in mammals relies on functional cooperation of different organs for the survival of the whole organism. The kidneys play a critical part in this process through the maintenance of fluid volume and composition homeostasis, which enables other organs to fulfil their tasks. The renal endothelium exhibits phenotypic and molecular traits that distinguish it from endothelia of other organs. Moreover, the adult kidney vasculature comprises diverse populations of mostly quiescent, but not metabolically inactive, endothelial cells (ECs) that reside within the kidney glomeruli, cortex and medulla. Each of these populations supports specific functions, for example, in the filtration of blood plasma, the reabsorption and secretion of water and solutes, and the concentration of urine. Transcriptional profiling of these diverse EC populations suggests they have adapted to local microenvironmental conditions (hypoxia, shear stress, hyperosmolarity), enabling them to support kidney functions. Exposure of ECs to microenvironment-derived angiogenic factors affects their metabolism, and sustains kidney development and homeostasis, whereas EC-derived angiocrine factors preserve distinct microenvironment niches. In the context of kidney disease, renal ECs show alteration in their metabolism and phenotype in response to pathological changes in the local microenvironment, further promoting kidney dysfunction. Understanding the diversity and specialization of kidney ECs could provide new avenues for the treatment of kidney diseases and kidney regeneration.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
