

Anterior gradient 2 increases long-chain fatty acid uptake via stabilizing FABP1 and facilitates lipid accumulation
- PMID: 33767592
- PMCID: PMC7975708
- DOI: 10.7150/ijbs.57099
Anterior gradient 2 increases long-chain fatty acid uptake via stabilizing FABP1 and facilitates lipid accumulation
Abstract
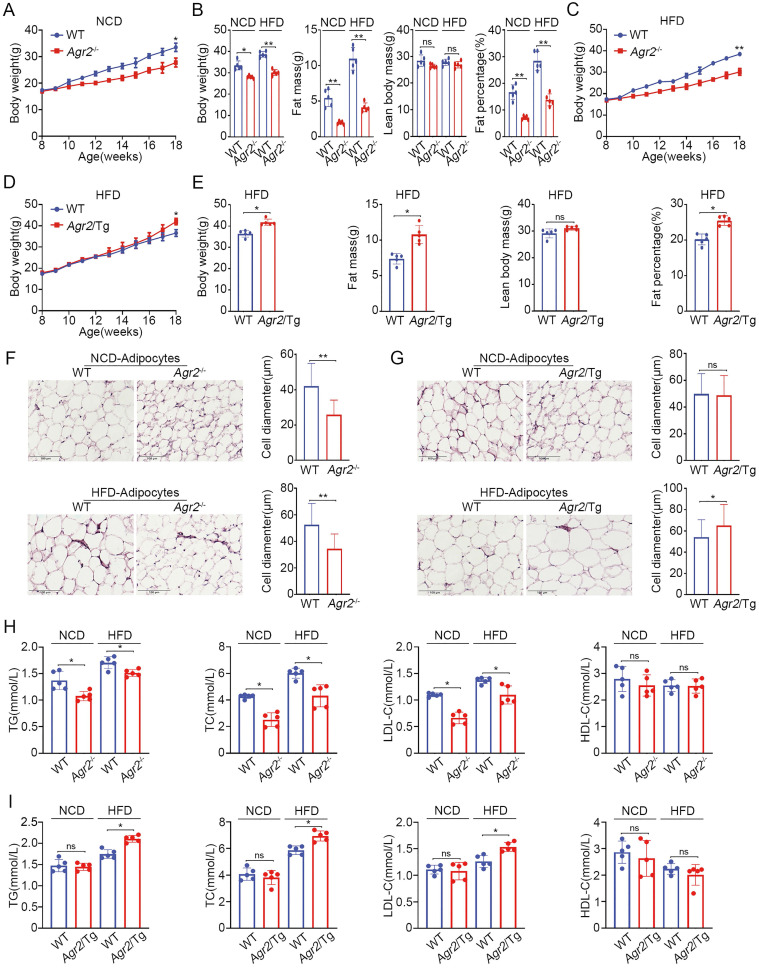
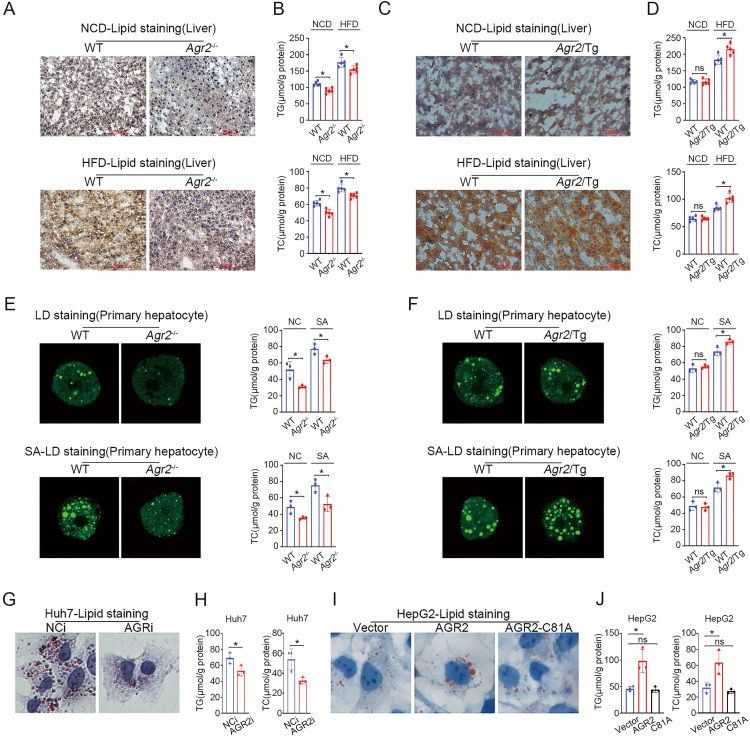
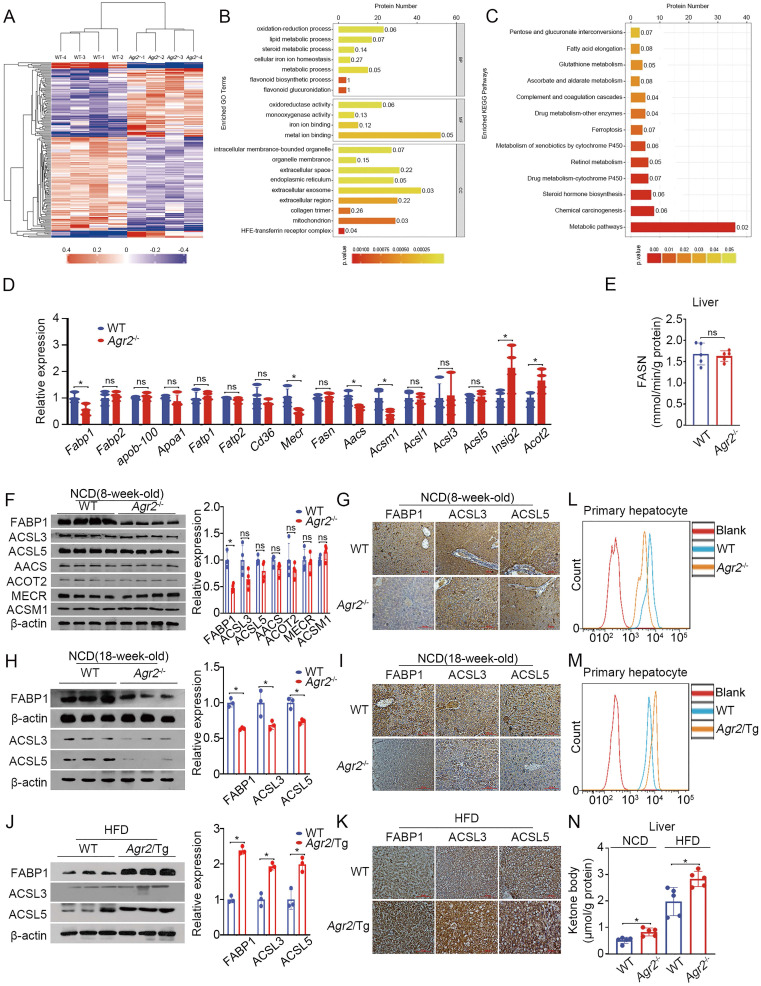
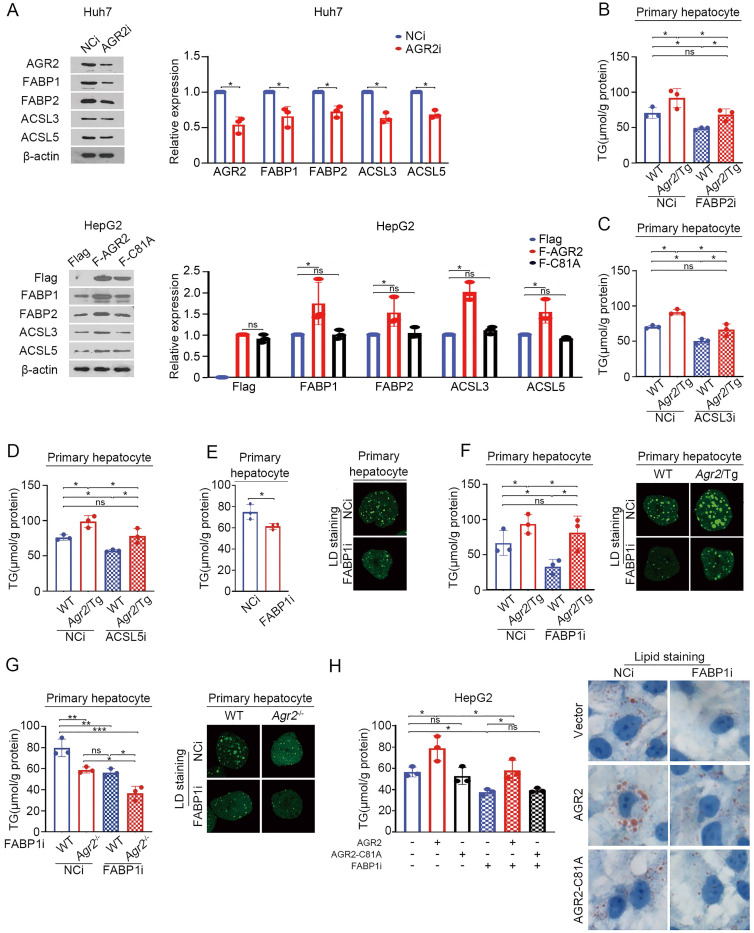
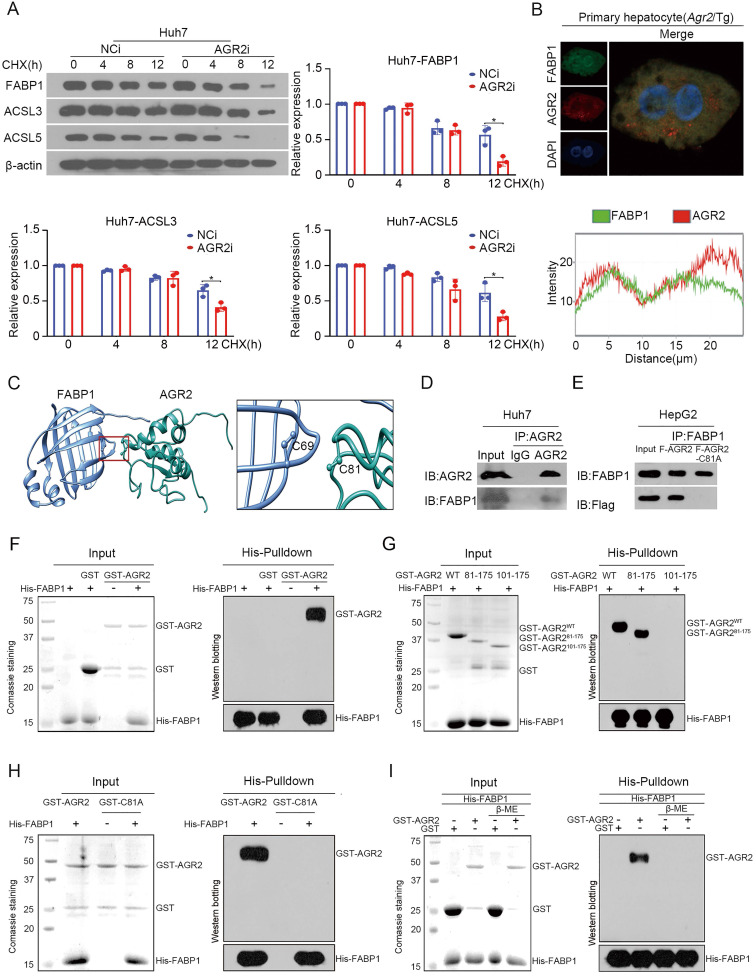
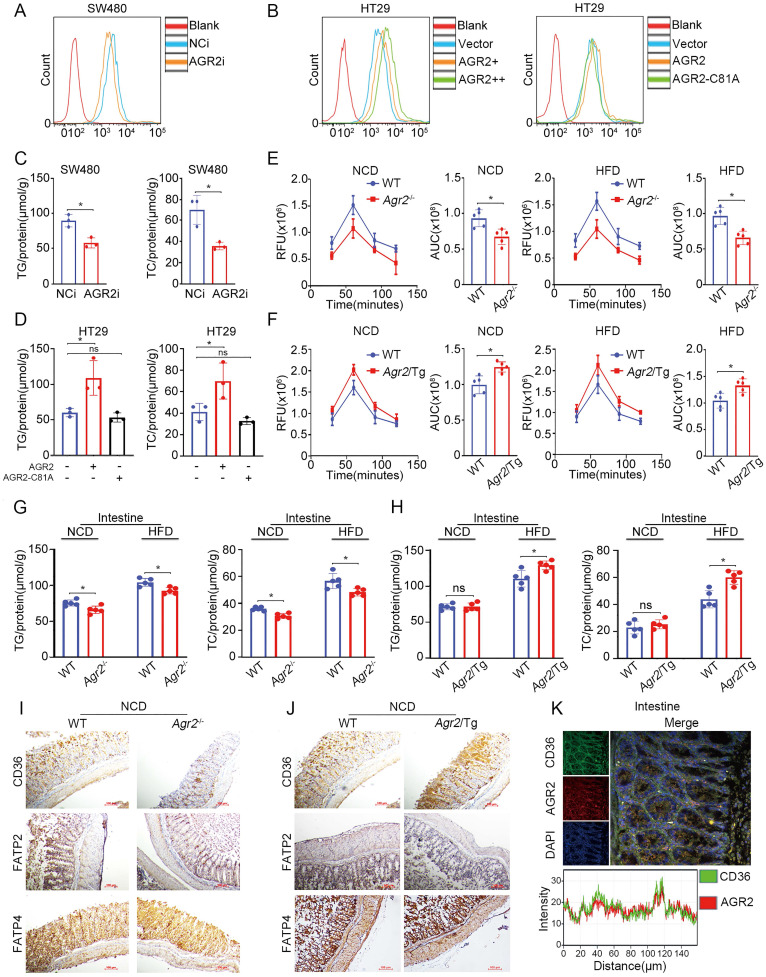
Anterior gradient 2 (AGR2), a protein disulfide isomerase (PDI), is a well-established oncogene. Here, we found that Agr2-/- mice had a decreased fat mass and hepatic and serum lipid levels compared with their wild-type littermates after fasting, and exhibited reduced high-fat diet (HFD)-induced fat accumulation. Transgenic mice overexpressing AGR2 (Agr2/Tg) readily gained fat weight on a HFD but not a normal diet. Proteomic analysis of hepatic samples from Agr2-/- mice revealed that depletion of AGR2 impaired long-chain fatty acid uptake and activation but did not affect de novo hepatic lipogenesis. Further investigations led to the identification of several effector substrates, particularly fatty acid binding protein-1 (FABP1) as essential for the AGR2-mediated effects. AGR2 was coexpressed with FABP1, and knockdown of AGR2 resulted in a reduction in FABP1 stability. Physical interactions of AGR2 and FABP1 depended on the PDI motif in AGR2 and the formation of a disulfide bond between these two proteins. Overexpression of AGR2 but not a mutant AGR2 protein lacking PDI activity suppressed lipid accumulation in cells lacking FABP1. Moreover, AGR2 deficiency significantly reduced fatty acid absorption in the intestine, which might be resulted from decreased fatty acid transporter CD36 in mice. These findings demonstrated a novel role of AGR2 in fatty-acid uptake and activation in both the liver and intestine, which contributed to the AGR2-mediated lipid accumulation, suggesting that AGR2 is an important regulator of whole-body lipid metabolism and down-regulation of AGR2 may antagonize the development of obesity.
Keywords: AGR2; FABP1; chaperone; intestine; lipid metabolism; liver.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures
Similar articles
-
Ablating both Fabp1 and Scp2/Scpx (TKO) induces hepatic phospholipid and cholesterol accumulation in high fat-fed mice.Biochim Biophys Acta Mol Cell Biol Lipids. 2018 Mar;1863(3):323-338. doi: 10.1016/j.bbalip.2017.12.013. Epub 2018 Jan 4. Biochim Biophys Acta Mol Cell Biol Lipids. 2018. PMID: 29307784 Free PMC article.
-
Intestinal peroxisome proliferator-activated receptor α-fatty acid-binding protein 1 axis modulates nonalcoholic steatohepatitis.Hepatology. 2023 Jan 1;77(1):239-255. doi: 10.1002/hep.32538. Epub 2022 May 15. Hepatology. 2023. PMID: 35460276 Free PMC article.
-
Long-term exercise prevents hepatic steatosis: a novel role of FABP1 in regulation of autophagy-lysosomal machinery.FASEB J. 2019 Nov;33(11):11870-11883. doi: 10.1096/fj.201900812R. Epub 2019 Jul 31. FASEB J. 2019. PMID: 31366243 Free PMC article.
-
Liver fatty acid-binding protein and obesity.J Nutr Biochem. 2010 Nov;21(11):1015-32. doi: 10.1016/j.jnutbio.2010.01.005. J Nutr Biochem. 2010. PMID: 20537520 Free PMC article. Review.
-
The anterior gradient-2 interactome.Am J Physiol Cell Physiol. 2020 Jan 1;318(1):C40-C47. doi: 10.1152/ajpcell.00532.2018. Epub 2019 Oct 23. Am J Physiol Cell Physiol. 2020. PMID: 31644305 Review.
Cited by
-
AGR2-induced cholesterol synthesis drives lovastatin resistance that is overcome by combination therapy with allicin.Acta Pharmacol Sin. 2022 Nov;43(11):2905-2916. doi: 10.1038/s41401-022-00909-3. Epub 2022 Apr 22. Acta Pharmacol Sin. 2022. PMID: 35459869 Free PMC article.
-
Gastric cancer adapts high lipid microenvironment via suppressing PPARG-FABP1 axis after arriving in the lymph node.Redox Biol. 2025 Jul 17;85:103759. doi: 10.1016/j.redox.2025.103759. Online ahead of print. Redox Biol. 2025. PMID: 40694956 Free PMC article.
-
AGR2-mediated cell-cell communication controls the antiviral immune response by promoting the thiol oxidation of TRAF3.Redox Biol. 2025 May;82:103581. doi: 10.1016/j.redox.2025.103581. Epub 2025 Mar 4. Redox Biol. 2025. PMID: 40085973 Free PMC article.
-
AGR2: The Covert Driver and New Dawn of Hepatobiliary and Pancreatic Cancer Treatment.Biomolecules. 2024 Jun 23;14(7):743. doi: 10.3390/biom14070743. Biomolecules. 2024. PMID: 39062458 Free PMC article. Review.
-
Gene Dynamics in Maternal Nutrition: The Role of Chromium in Mitigating Offspring Obesity.Biol Trace Elem Res. 2025 Jun 19. doi: 10.1007/s12011-025-04711-0. Online ahead of print. Biol Trace Elem Res. 2025. PMID: 40536644
References
-
- Yang L, Zhao D, Ren J, Yang J. Endoplasmic reticulum stress and protein quality control in diabetic cardiomyopathy. Biochimica et biophysica acta. 2015;1852:209–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
