

CD46 splice variant enhances translation of specific mRNAs linked to an aggressive tumor cell phenotype in bladder cancer
- PMID: 33767911
- PMCID: PMC7972933
- DOI: 10.1016/j.omtn.2021.02.019
CD46 splice variant enhances translation of specific mRNAs linked to an aggressive tumor cell phenotype in bladder cancer
Abstract
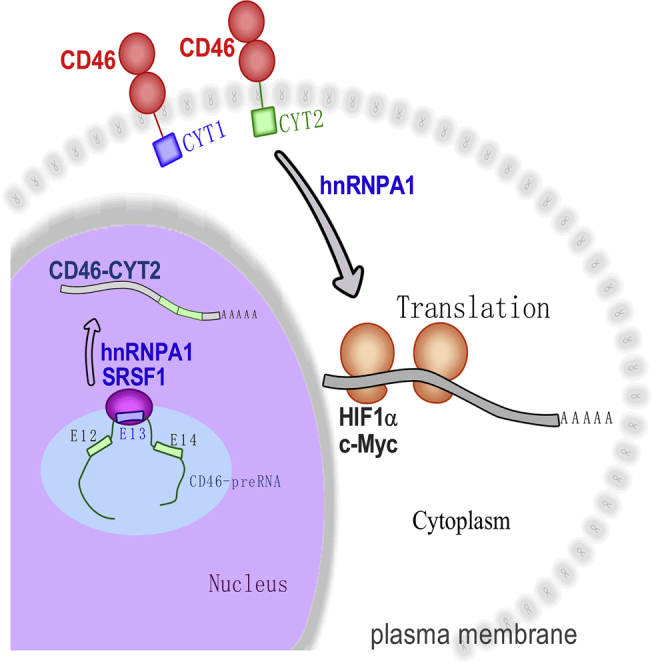
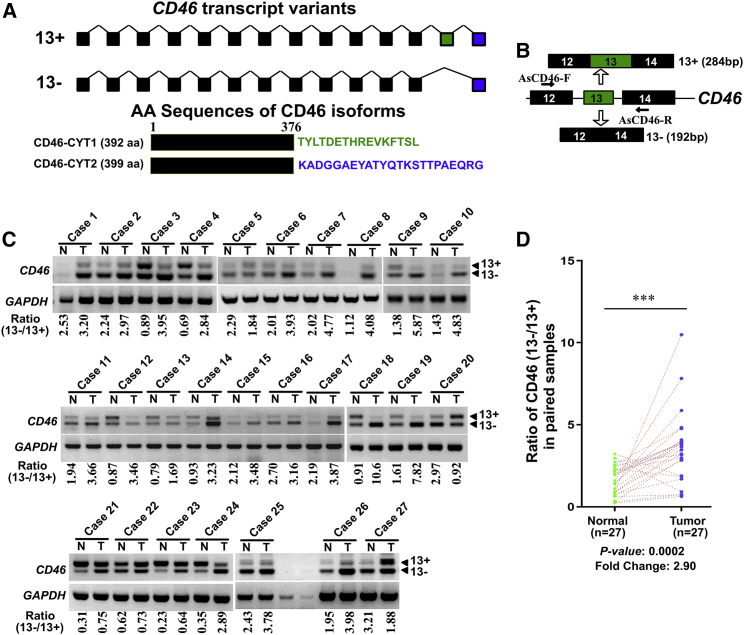
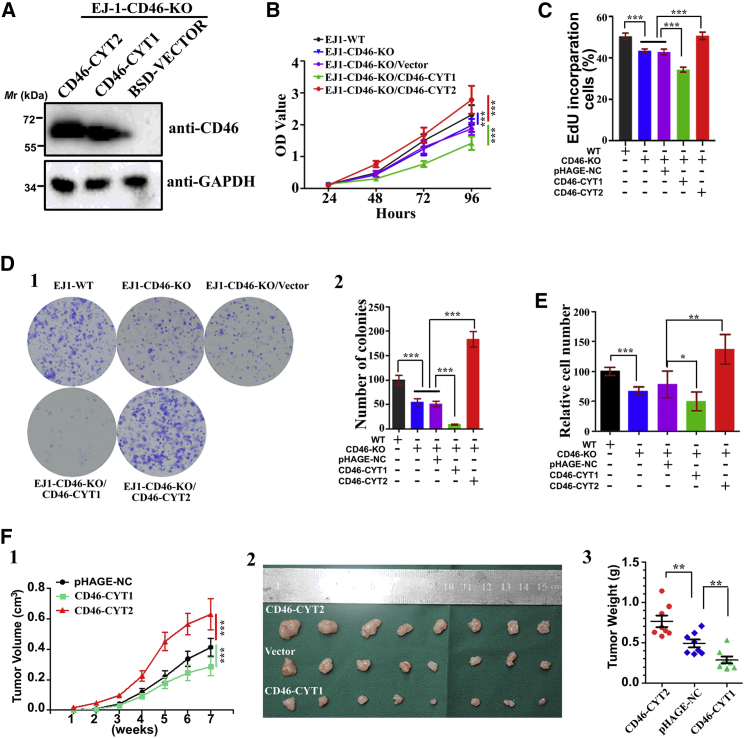
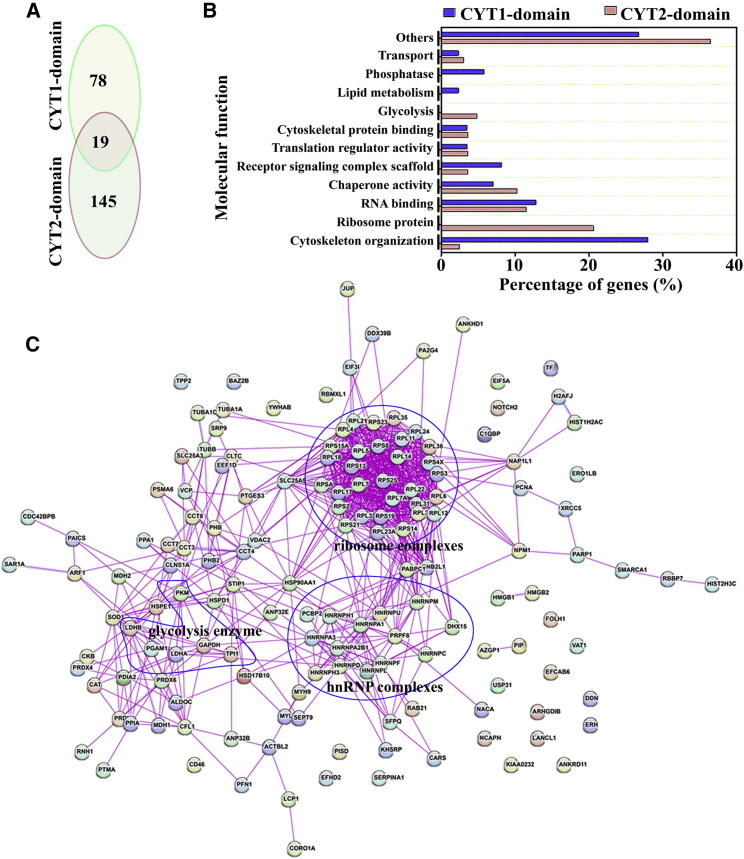
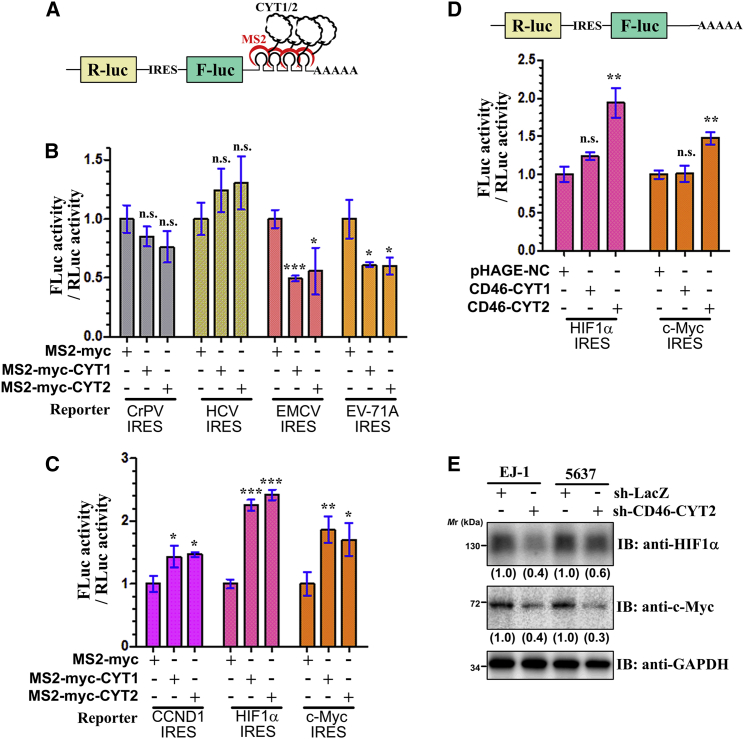
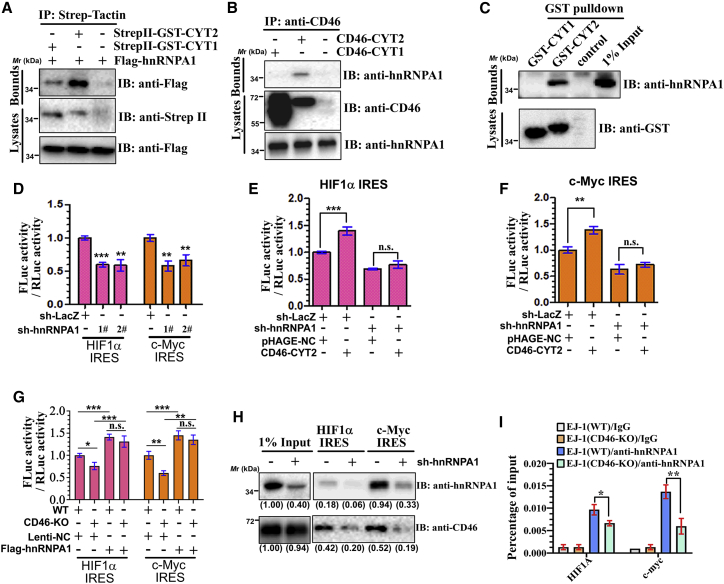
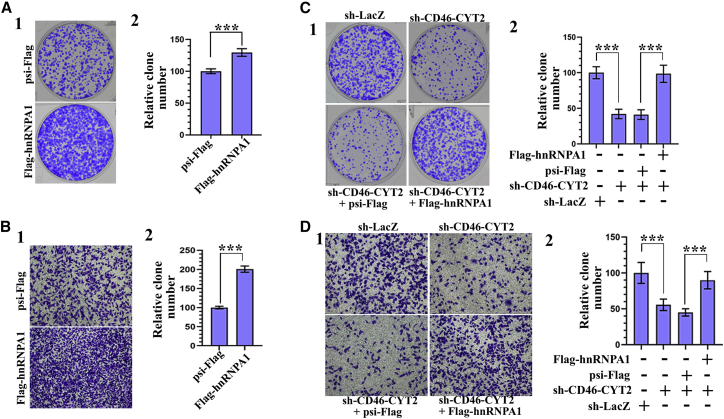
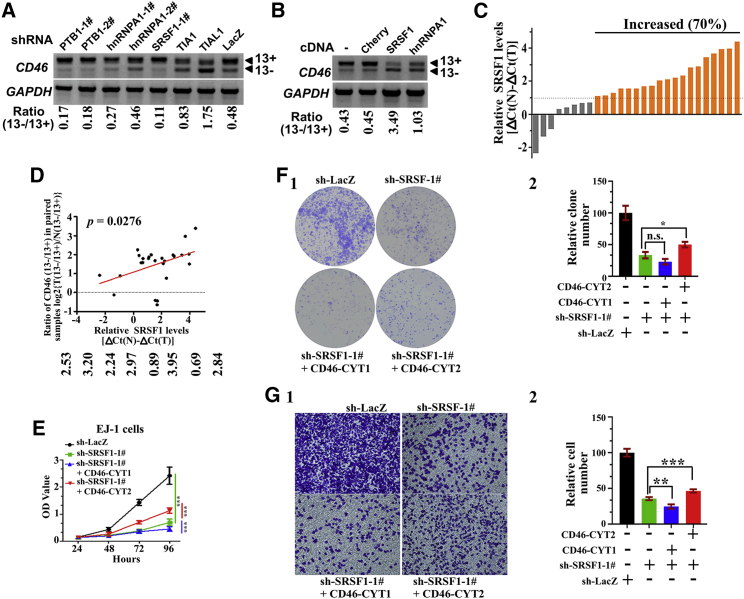
CD46 is well known to be involved in diverse biological processes. Although several splice variants of CD46 have been identified, little is known about the contribution of alternative splicing to its tumorigenic functions. In this study, we found that exclusion of CD46 exon 13 is significantly increased in bladder cancer (BCa) samples. In BCa cell lines, enforced expression of CD46-CYT2 (exon 13-skipping isoform) promoted, and CD46-CYT1 (exon 13-containing isoform) attenuated, cell growth, migration, and tumorigenicity in a xenograft model. We also applied interaction proteomics to identify exhaustively the complexes containing the CYT1 or CYT2 domain in EJ-1 cells. 320 proteins were identified that interact with the CYT1 and/or CYT2 domain, and most of them are new interactors. Using an internal ribosome entry site (IRES)-dependent reporter system, we established that CD46 could regulate mRNA translation through an interaction with the translation machinery. We also identified heterogeneous nuclear ribonucleoprotein (hnRNP)A1 as a novel CYT2 binding partner, and this interaction facilitates the interaction of hnRNPA1 with IRES RNA to promote IRES-dependent translation of HIF1a and c-Myc. Strikingly, the splicing factor SRSF1 is highly correlated with CD46 exon 13 exclusion in clinical BCa samples. Taken together, our findings contribute to understanding the role of CD46 in BCa development.
Keywords: CD46; bladder cancer; hnRNPA1; translation; tumorigenesis.
© 2021 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Presenilin/gamma-secretase cleaves CD46 in response to Neisseria infection.J Immunol. 2010 Jan 15;184(2):694-701. doi: 10.4049/jimmunol.0900522. Epub 2009 Dec 16. J Immunol. 2010. PMID: 20018629 Free PMC article.
-
High expression of membrane cofactor protein of complement (CD46) in human leukaemia cell lines: implication of an alternatively spliced form containing the STA domain in CD46 up-regulation.Scand J Immunol. 1995 Dec;42(6):581-90. doi: 10.1111/j.1365-3083.1995.tb03700.x. Scand J Immunol. 1995. PMID: 8552981
-
PRMT5 regulates IRES-dependent translation via methylation of hnRNP A1.Nucleic Acids Res. 2017 May 5;45(8):4359-4369. doi: 10.1093/nar/gkw1367. Nucleic Acids Res. 2017. PMID: 28115626 Free PMC article.
-
RBMX suppresses tumorigenicity and progression of bladder cancer by interacting with the hnRNP A1 protein to regulate PKM alternative splicing.Oncogene. 2021 Apr;40(15):2635-2650. doi: 10.1038/s41388-021-01666-z. Epub 2021 Feb 9. Oncogene. 2021. PMID: 33564070 Free PMC article.
-
hnRNP A1 in RNA metabolism regulation and as a potential therapeutic target.Front Pharmacol. 2022 Oct 21;13:986409. doi: 10.3389/fphar.2022.986409. eCollection 2022. Front Pharmacol. 2022. PMID: 36339596 Free PMC article. Review.
Cited by
-
Comprehensive Analysis of Prognostic Alternative Splicing Signature Reveals Recurrence Predictor for Papillary Thyroid Cancer.Front Oncol. 2021 Oct 13;11:705929. doi: 10.3389/fonc.2021.705929. eCollection 2021. Front Oncol. 2021. PMID: 34722250 Free PMC article.
-
OncoSplicing 3.0: an updated database for identifying RBPs regulating alternative splicing events in cancers.Nucleic Acids Res. 2025 Jan 6;53(D1):D1460-D1466. doi: 10.1093/nar/gkae1098. Nucleic Acids Res. 2025. PMID: 39558172 Free PMC article.
-
Complement System and the Kidney: Its Role in Renal Diseases, Kidney Transplantation and Renal Cell Carcinoma.Int J Mol Sci. 2023 Nov 20;24(22):16515. doi: 10.3390/ijms242216515. Int J Mol Sci. 2023. PMID: 38003705 Free PMC article. Review.
-
Complement regulatory protein CD46 promotes bladder cancer metastasis through activation of MMP9.Int J Oncol. 2024 Jul;65(1):71. doi: 10.3892/ijo.2024.5659. Epub 2024 Jun 7. Int J Oncol. 2024. PMID: 38847230 Free PMC article.
-
Alternative splicing regulation and its therapeutic potential in bladder cancer.Front Oncol. 2024 Jul 26;14:1402350. doi: 10.3389/fonc.2024.1402350. eCollection 2024. Front Oncol. 2024. PMID: 39132499 Free PMC article. Review.
References
-
- Sveen A., Kilpinen S., Ruusulehto A., Lothe R.A., Skotheim R.I. Aberrant RNA splicing in cancer; expression changes and driver mutations of splicing factor genes. Oncogene. 2016;35:2413–2427. - PubMed
-
- Climente-González H., Porta-Pardo E., Godzik A., Eyras E. The functional impact of alternative splicing in cancer. Cell Rep. 2017;20:2215–2226. - PubMed
-
- Siegfried Z., Karni R. The role of alternative splicing in cancer drug resistance. Curr. Opin. Genet. Dev. 2018;48:16–21. - PubMed
-
- Yamamoto H., Fara A.F., Dasgupta P., Kemper C. CD46: the “multitasker” of complement proteins. Int. J. Biochem. Cell Biol. 2013;45:2808–2820. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
