

Arabidopsis cryptochrome 1 controls photomorphogenesis through regulation of H2A.Z deposition
- PMID: 33768238
- PMCID: PMC8290288
- DOI: 10.1093/plcell/koab091
Arabidopsis cryptochrome 1 controls photomorphogenesis through regulation of H2A.Z deposition
Abstract
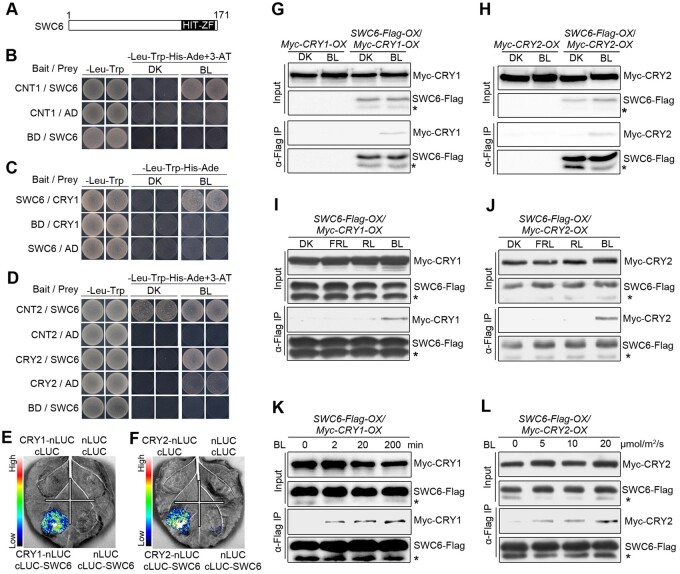
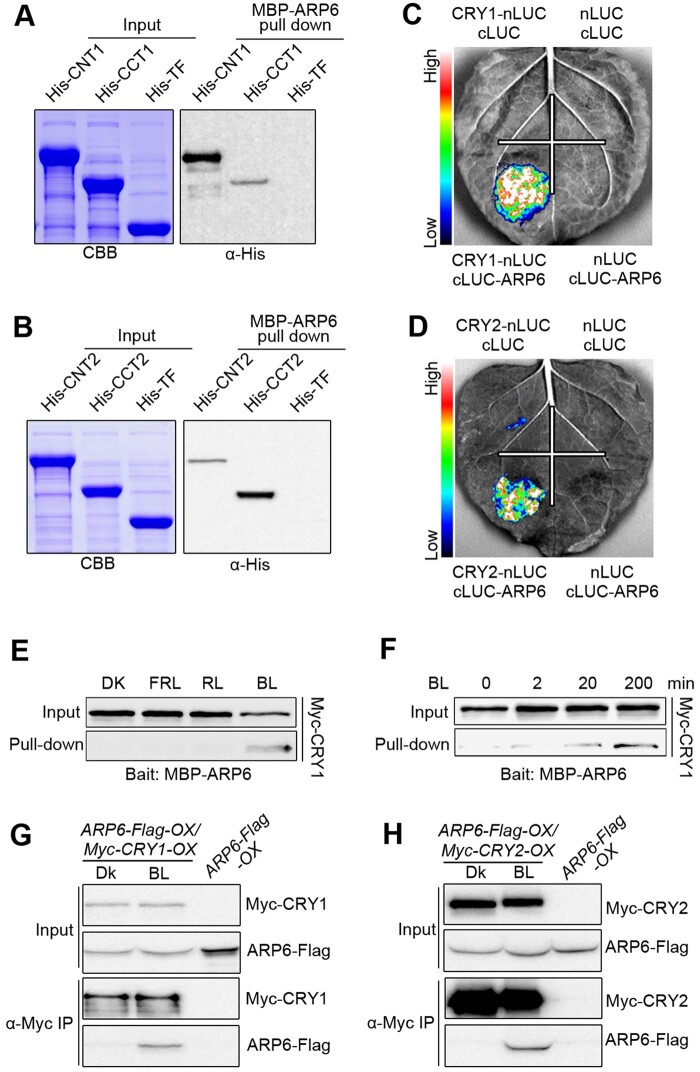
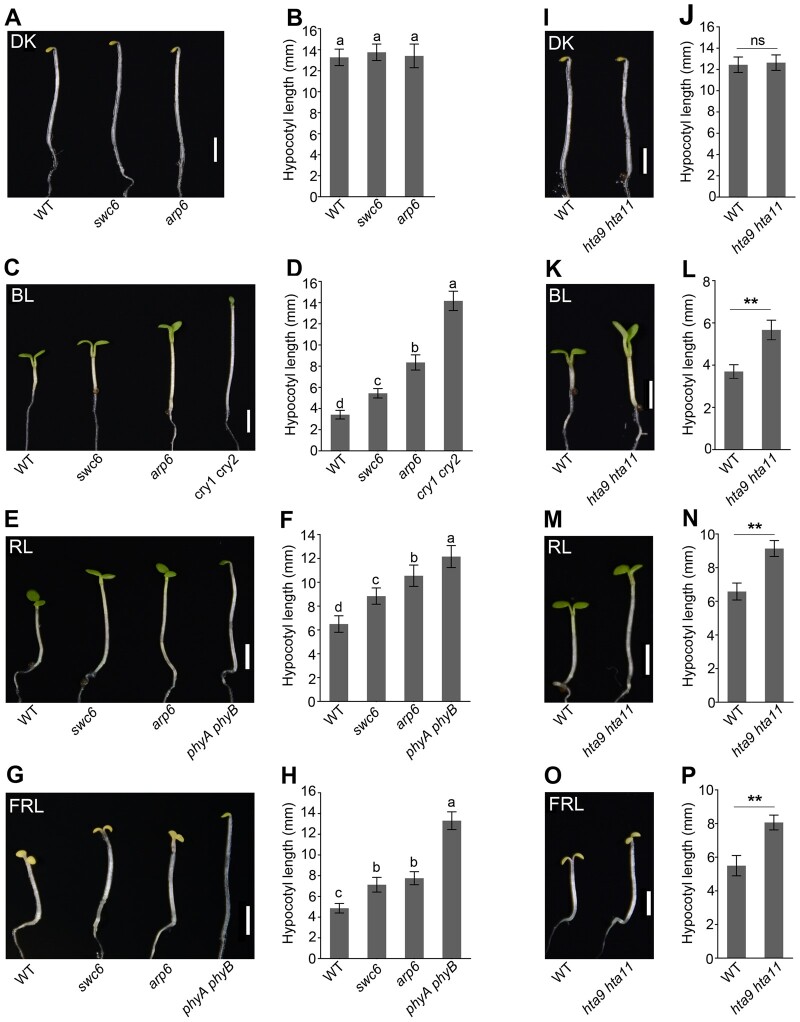
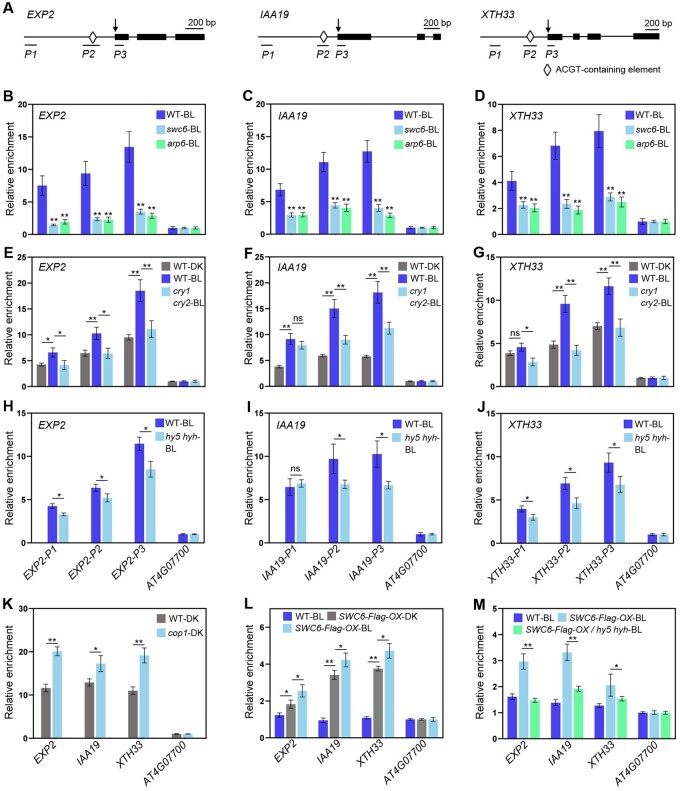
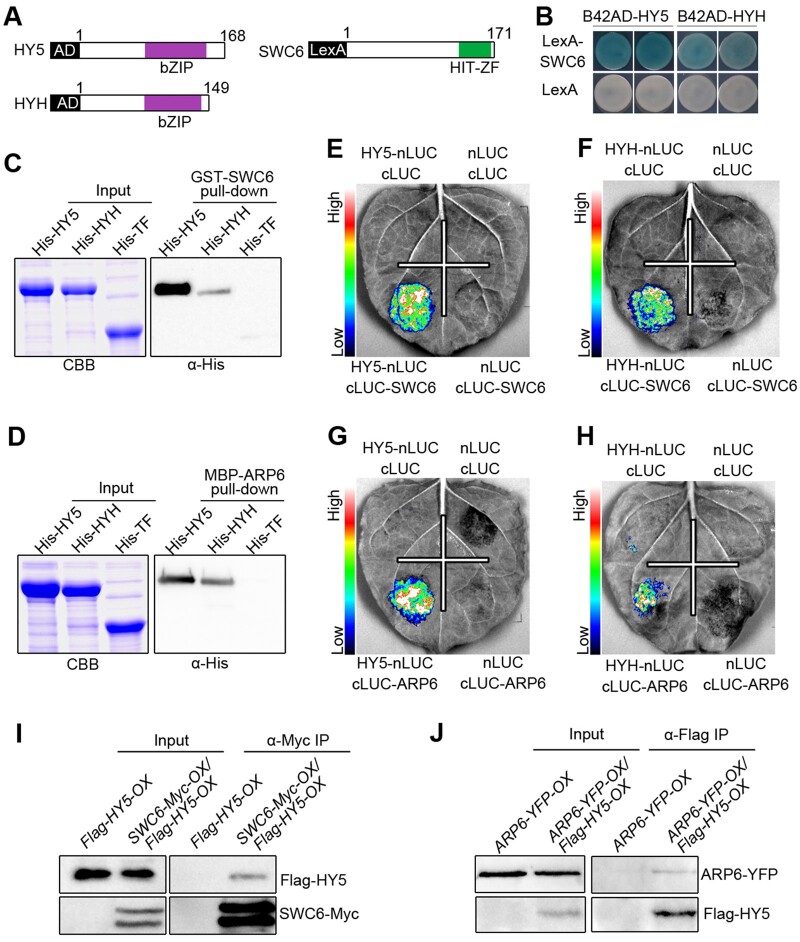
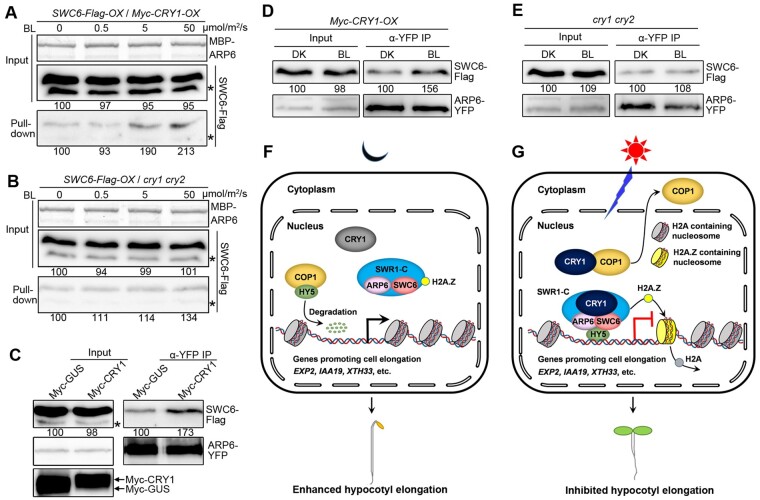
Light is a key environmental cue that fundamentally regulates plant growth and development, which is mediated by the multiple photoreceptors including the blue light (BL) photoreceptor cryptochrome 1 (CRY1). The signaling mechanism of Arabidopsis thaliana CRY1 involves direct interactions with CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1)/SUPPRESSOR OF PHYA-105 1 and stabilization of COP1 substrate ELONGATED HYPOCOTYL 5 (HY5). H2A.Z is an evolutionarily conserved histone variant, which plays a critical role in transcriptional regulation through its deposition in chromatin catalyzed by SWR1 complex. Here we show that CRY1 physically interacts with SWC6 and ARP6, the SWR1 complex core subunits that are essential for mediating H2A.Z deposition, in a BL-dependent manner, and that BL-activated CRY1 enhances the interaction of SWC6 with ARP6. Moreover, HY5 physically interacts with SWC6 and ARP6 to direct the recruitment of SWR1 complex to HY5 target loci. Based on previous studies and our findings, we propose that CRY1 promotes H2A.Z deposition to regulate HY5 target gene expression and photomorphogenesis in BL through the enhancement of both SWR1 complex activity and HY5 recruitment of SWR1 complex to HY5 target loci, which is likely mediated by interactions of CRY1 with SWC6 and ARP6, and CRY1 stabilization of HY5, respectively.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures

References
-
- Ahmad M, Cashmore AR (1993) HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature 366: 162–166 - PubMed
-
- Ang LH, Chattopadhyay S, Wei N, Oyama T, Okada K, Batschauer A, Deng XW (1998) Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol Cell 1: 213–222 - PubMed
-
- Briggs WR, Christie JM (2002) Phototropins 1 and 2: versatile plant blue-light receptors. Trends Plant Sci 7: 204–210 - PubMed
-
- Bruggemann E, Handwerger K, Essex C, Storz G (1996) Analysis of fast neutron-generated mutants at the Arabidopsis thaliana HY4 locus. Plant J 10: 755–760 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
