

Structure of the EphB6 receptor ectodomain
- PMID: 33770085
- PMCID: PMC7997048
- DOI: 10.1371/journal.pone.0247335
Structure of the EphB6 receptor ectodomain
Abstract
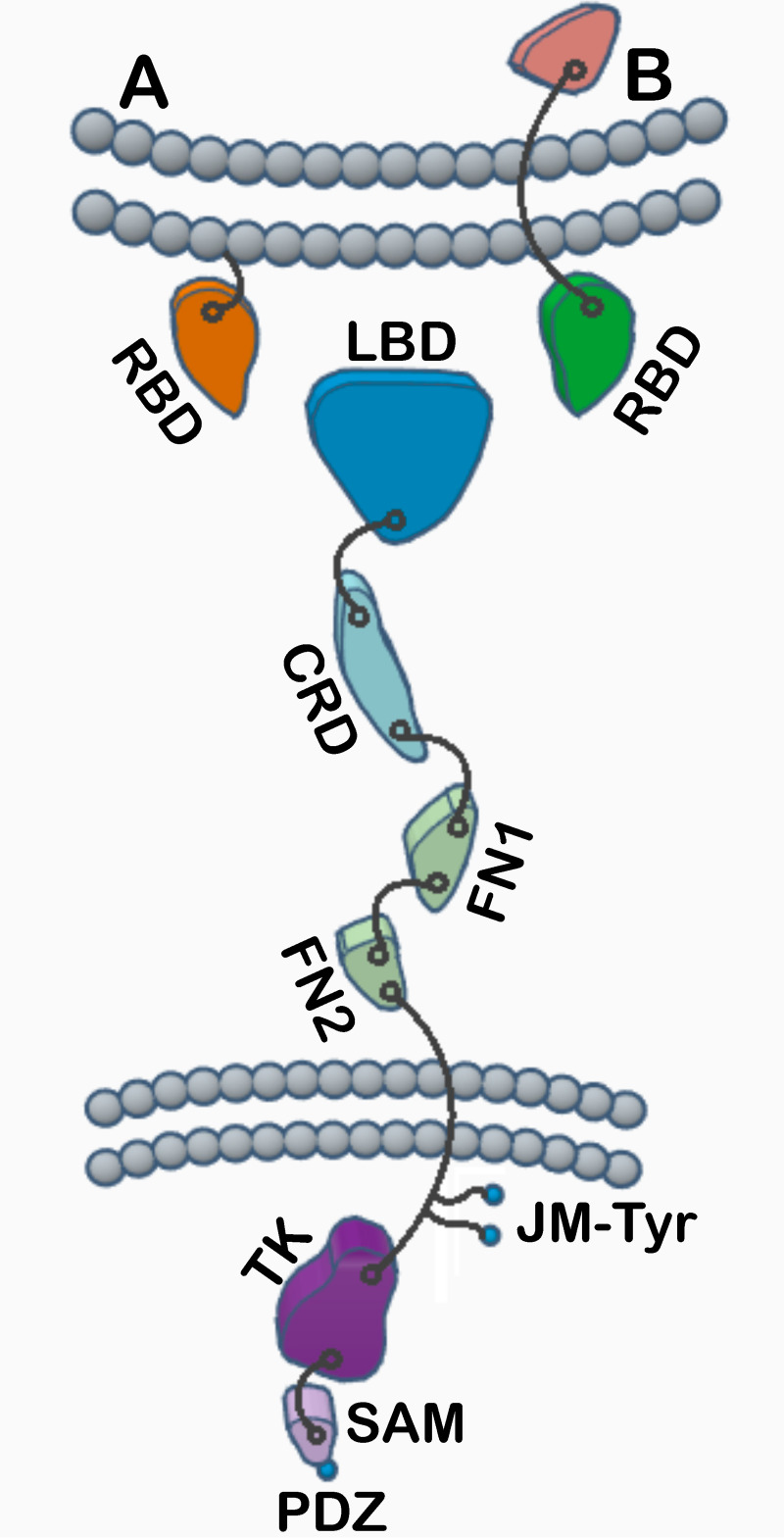
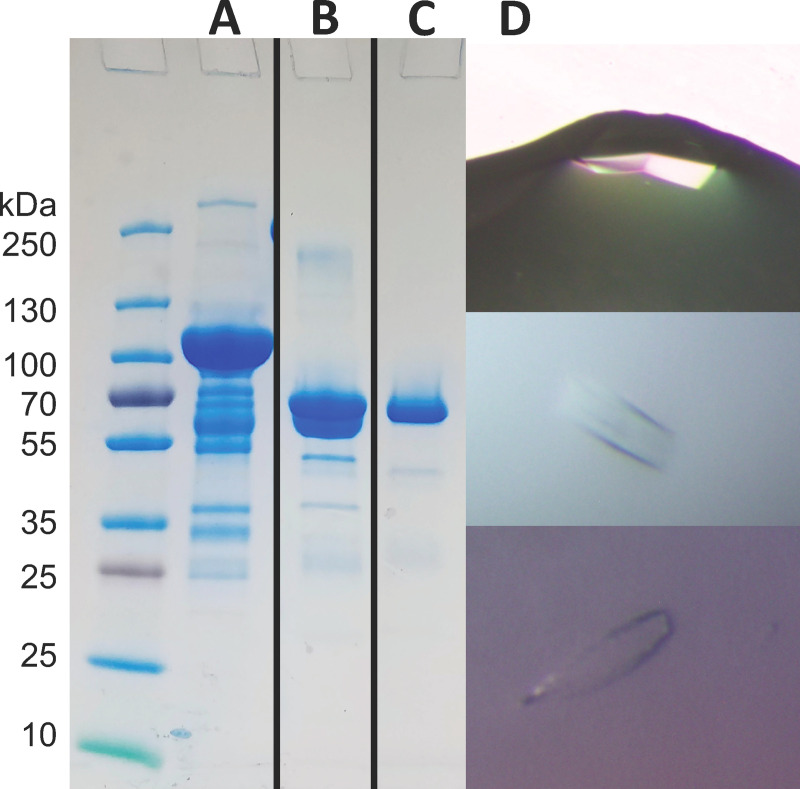
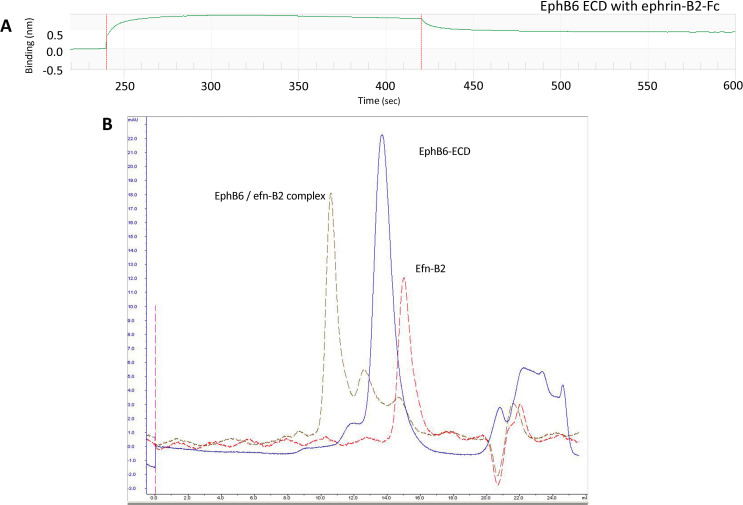
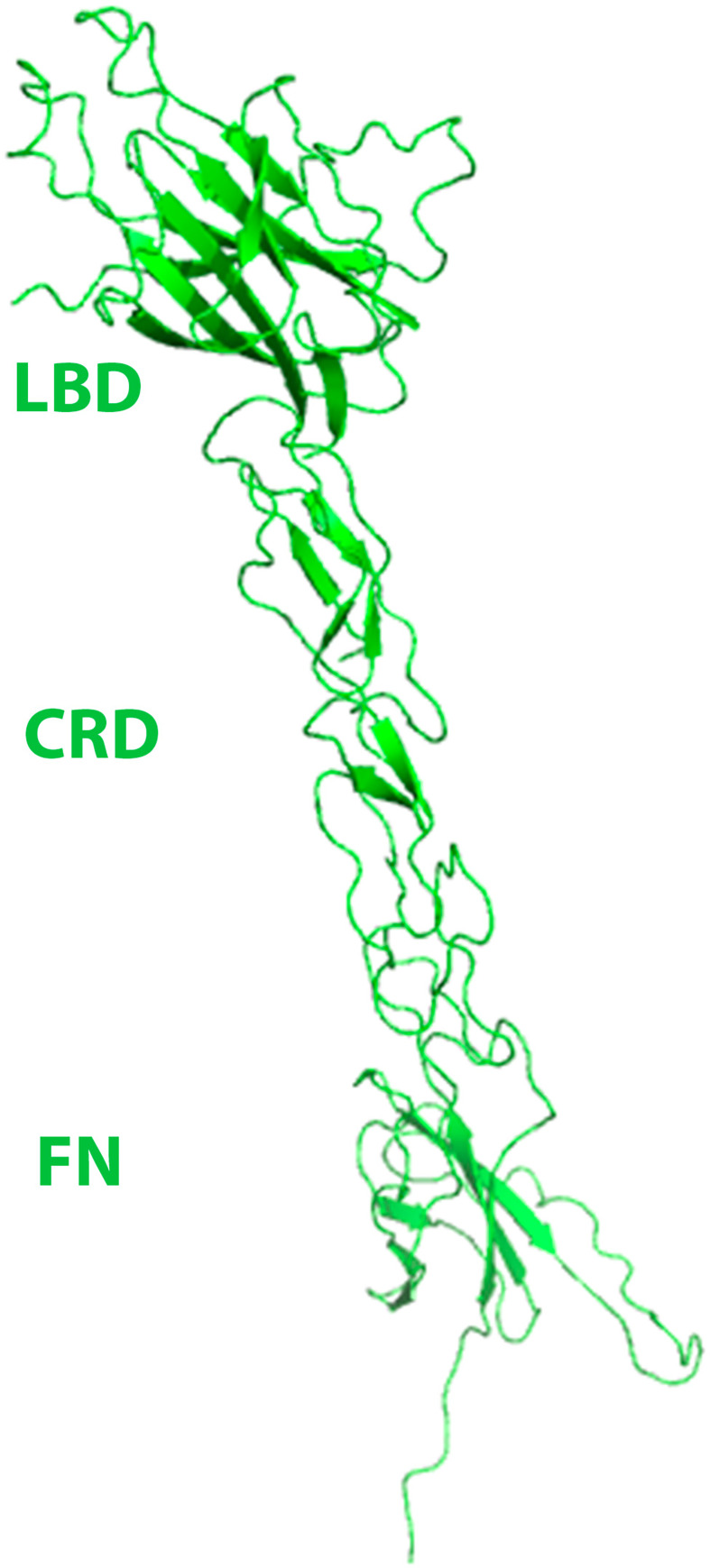
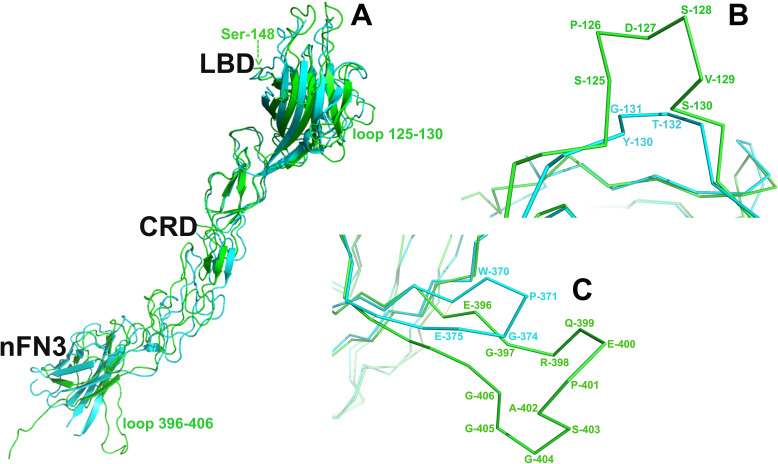
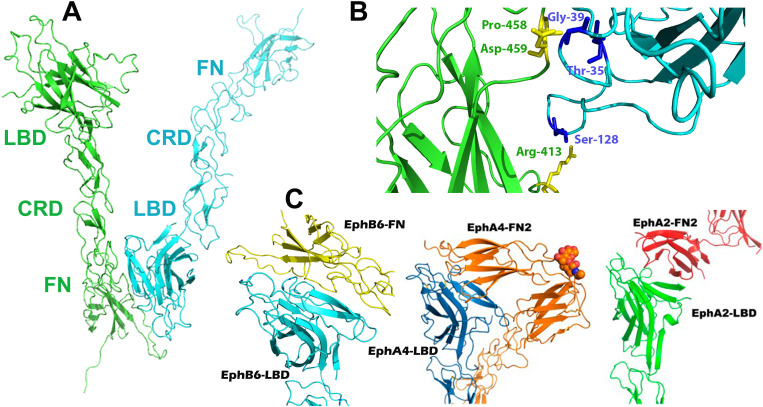
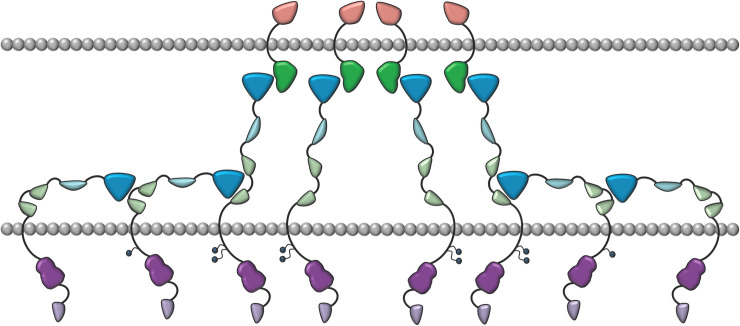
Eph receptors are the largest group amongst the receptor tyrosine kinases and are divided into two subgroups, A and B, based on ligand binding specificities and sequence conservation. Through ligand-induced and ligand-independent activities, Ephs play central roles in diverse biological processes, including embryo development, regulation of neuronal signaling, immune responses, vasculogenesis, as well as tumor initiation, progression, and metastasis. The Eph extracellular regions (ECDs) are constituted of multiple domains, and previous structural studies of the A class receptors revealed how they interact with ephrin ligands and simultaneously mediate Eph-Eph clustering necessary for biological activity. Specifically, EphA structures highlighted a model, where clustering of ligand-bound receptors relies on two distinct receptor/receptor interfaces. Interestingly, most unliganded A class receptors also form an additional, third interface, between the ligand binding domain (LBD) and the fibronectin III domain (FN3) of neighboring molecules. Structures of B-class Eph ECDs, on the other hand, have never been reported. To further our understanding of Eph receptor function, we crystallized the EphB6-ECD and determined its three-dimensional structure using X-ray crystallography. EphB6 has important functions in both normal physiology and human malignancies and is especially interesting because this atypical receptor innately lacks kinase activity and our understanding of the mechanism of action is still incomplete. Our structural data reveals the overall EphB6-ECD architecture and shows EphB6-LBD/FN3 interactions similar to those observed for the unliganded A class receptors, suggesting that these unusual interactions are of general importance to the Eph group. We also observe unique structural features, which likely reflect the atypical signaling properties of EphB6, namely the need of co-receptor(s) for this kinase-inactive Eph. These findings provide new valuable information on the structural organization and mechanism of action of the B-class Ephs, and specifically EphB6, which in the future will assist in identifying clinically relevant targets for cancer therapy.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
