

Malic enzyme 2 connects the Krebs cycle intermediate fumarate to mitochondrial biogenesis
- PMID: 33770508
- PMCID: PMC10472834
- DOI: 10.1016/j.cmet.2021.03.003
Malic enzyme 2 connects the Krebs cycle intermediate fumarate to mitochondrial biogenesis
Abstract
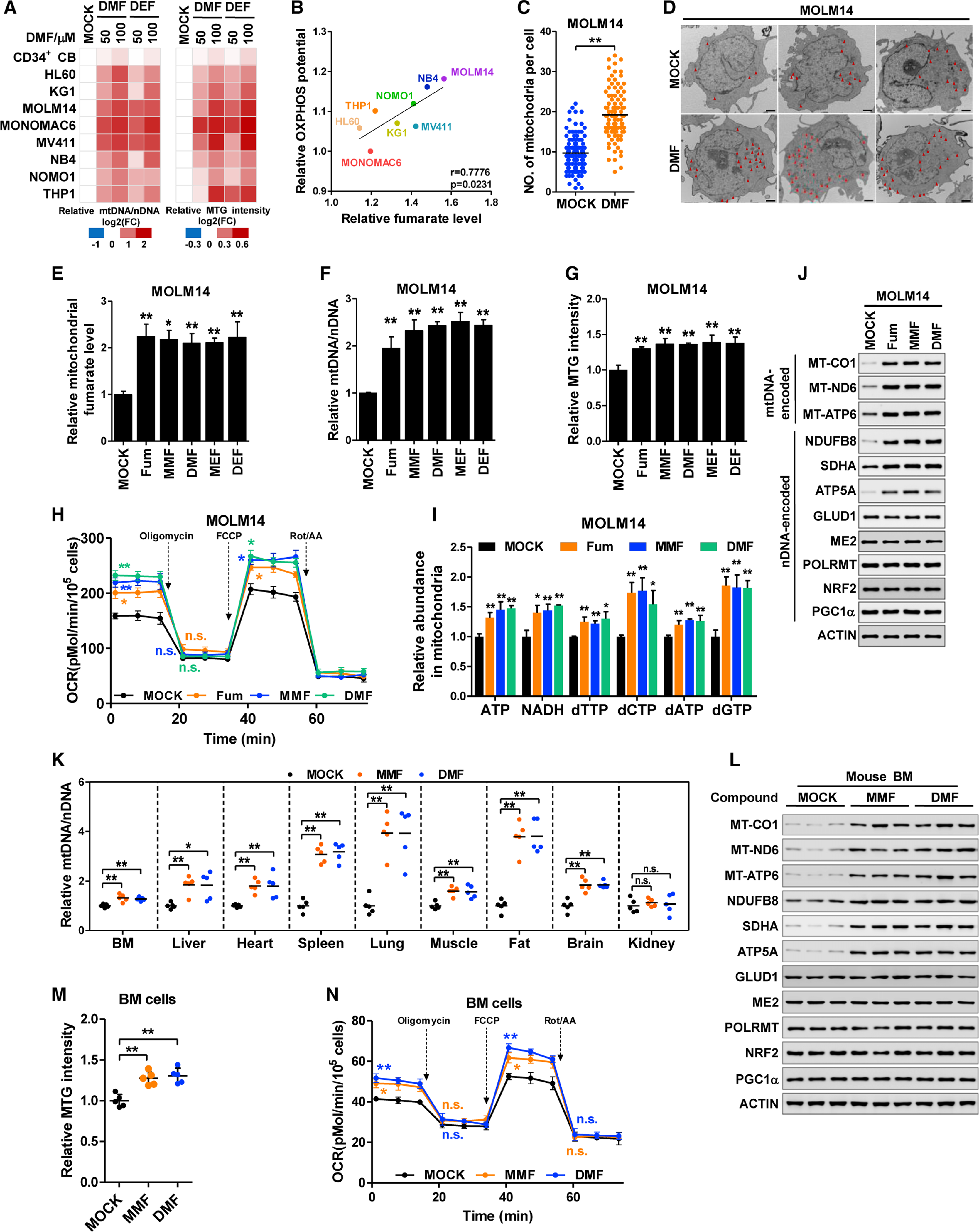
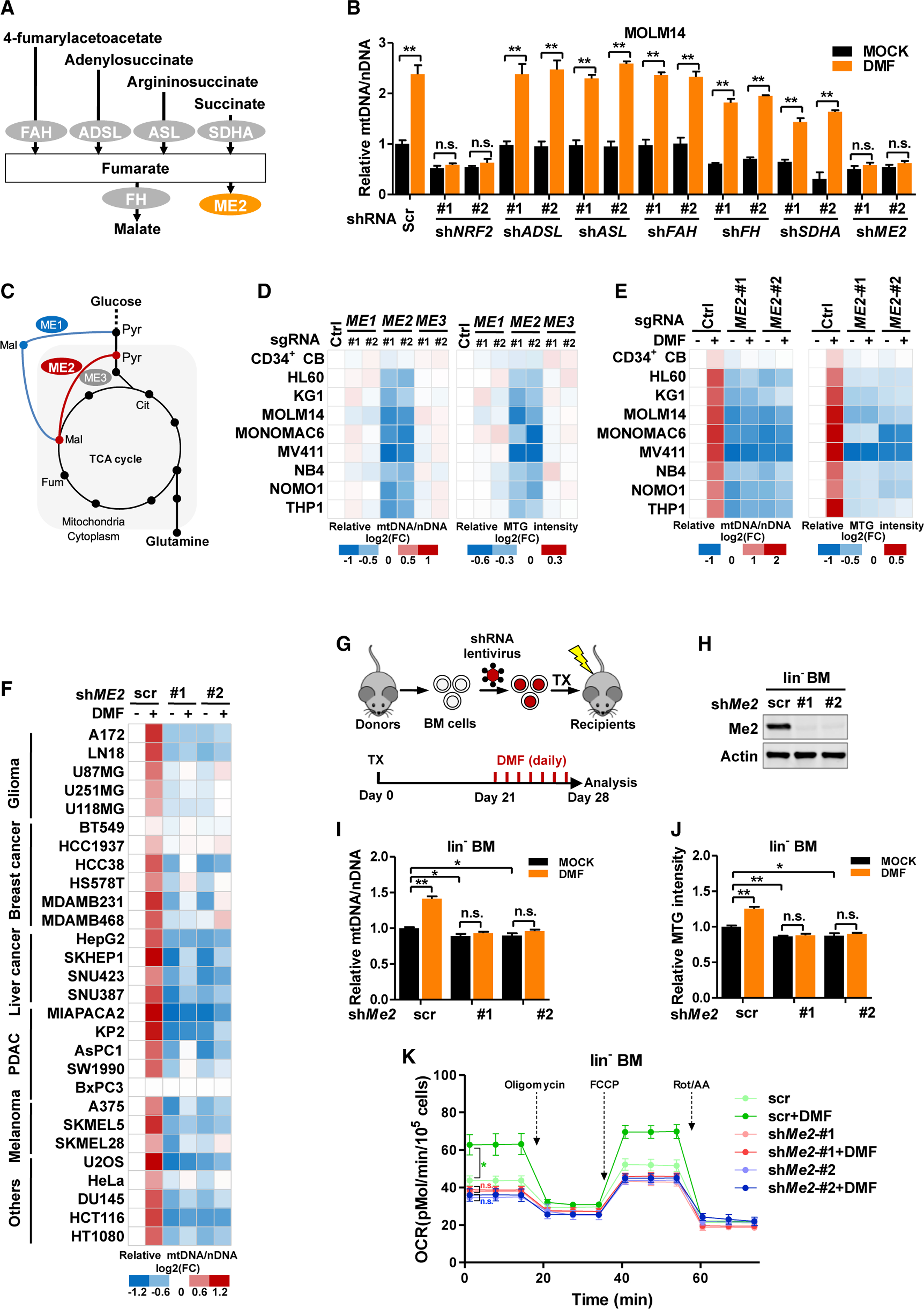
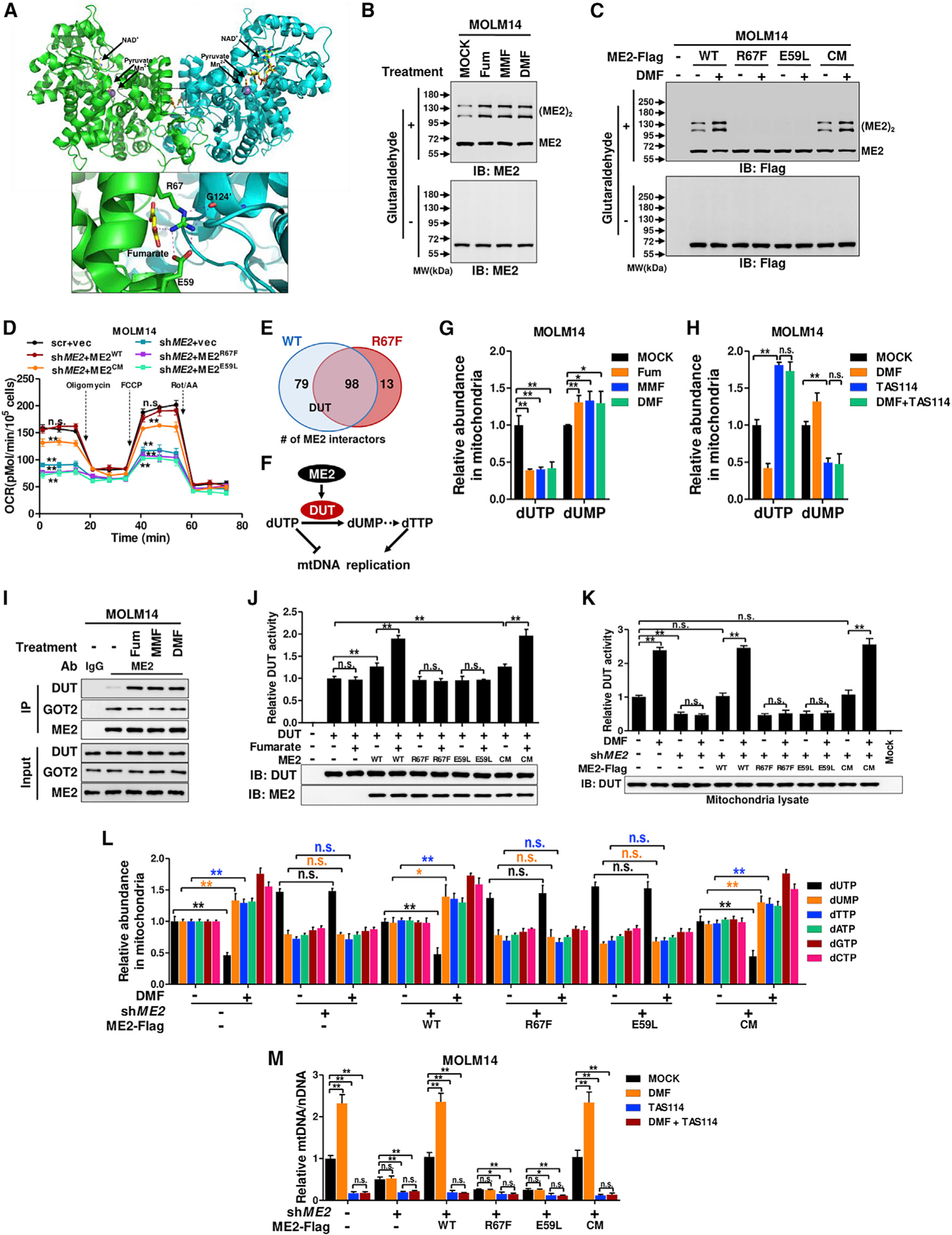
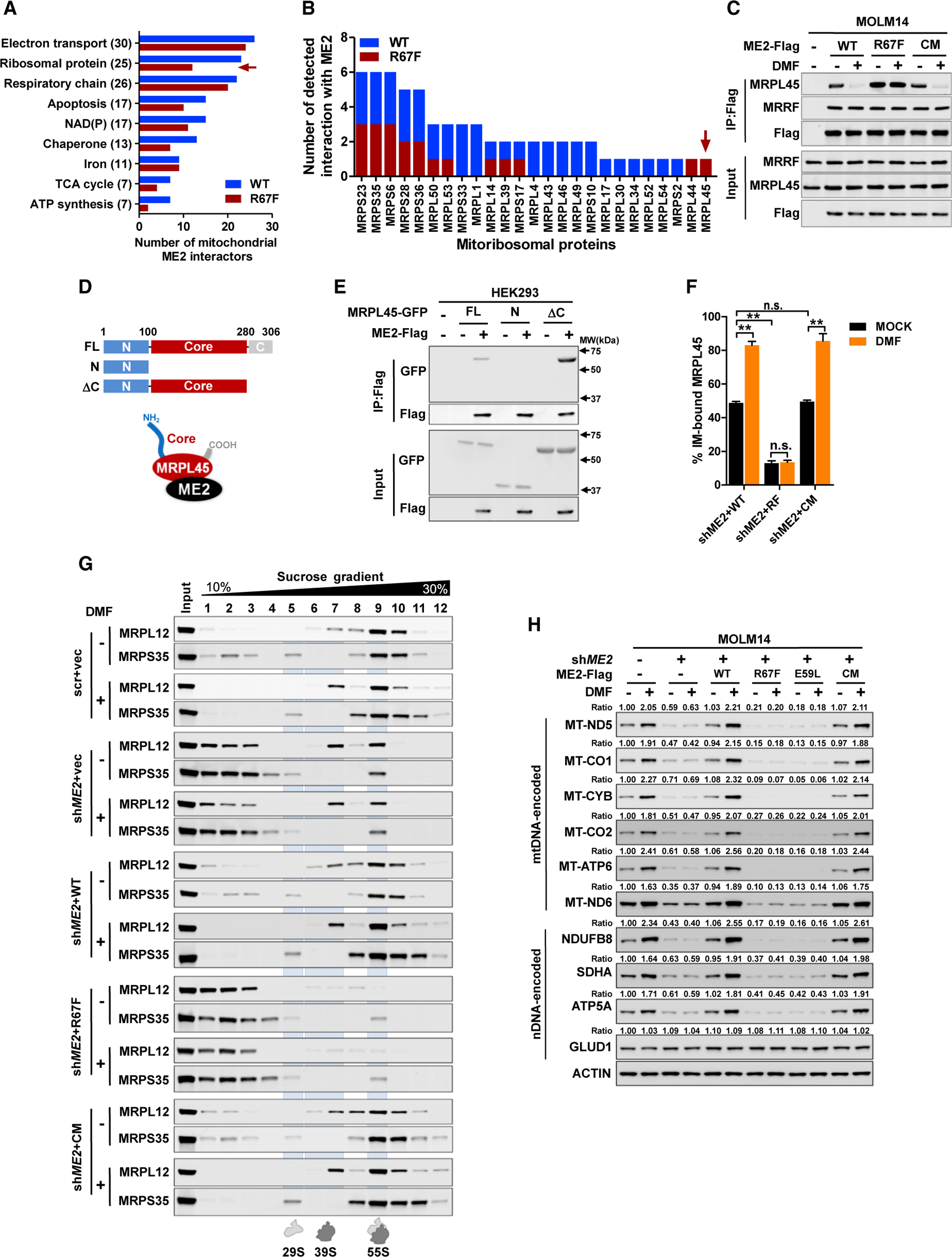
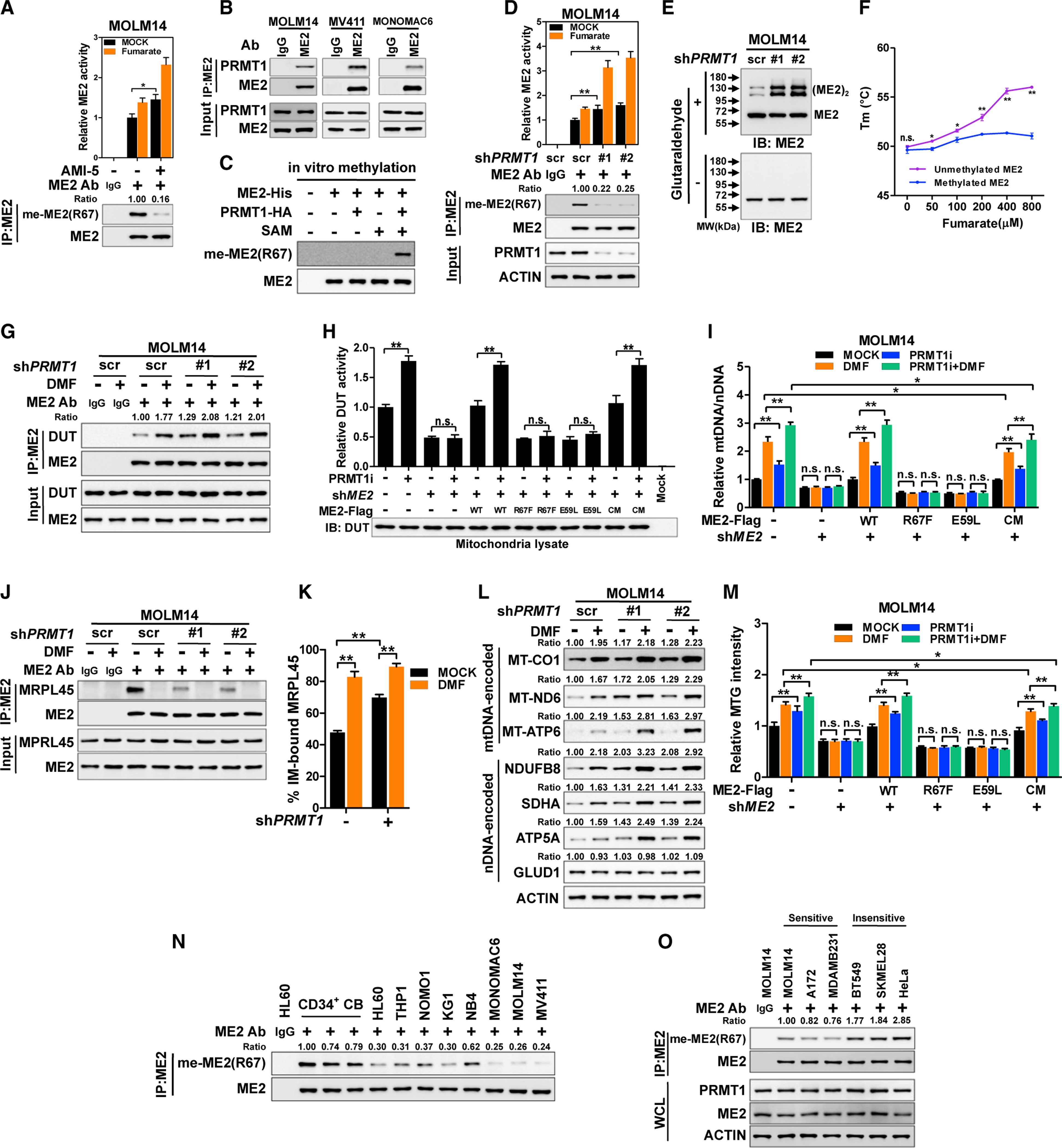
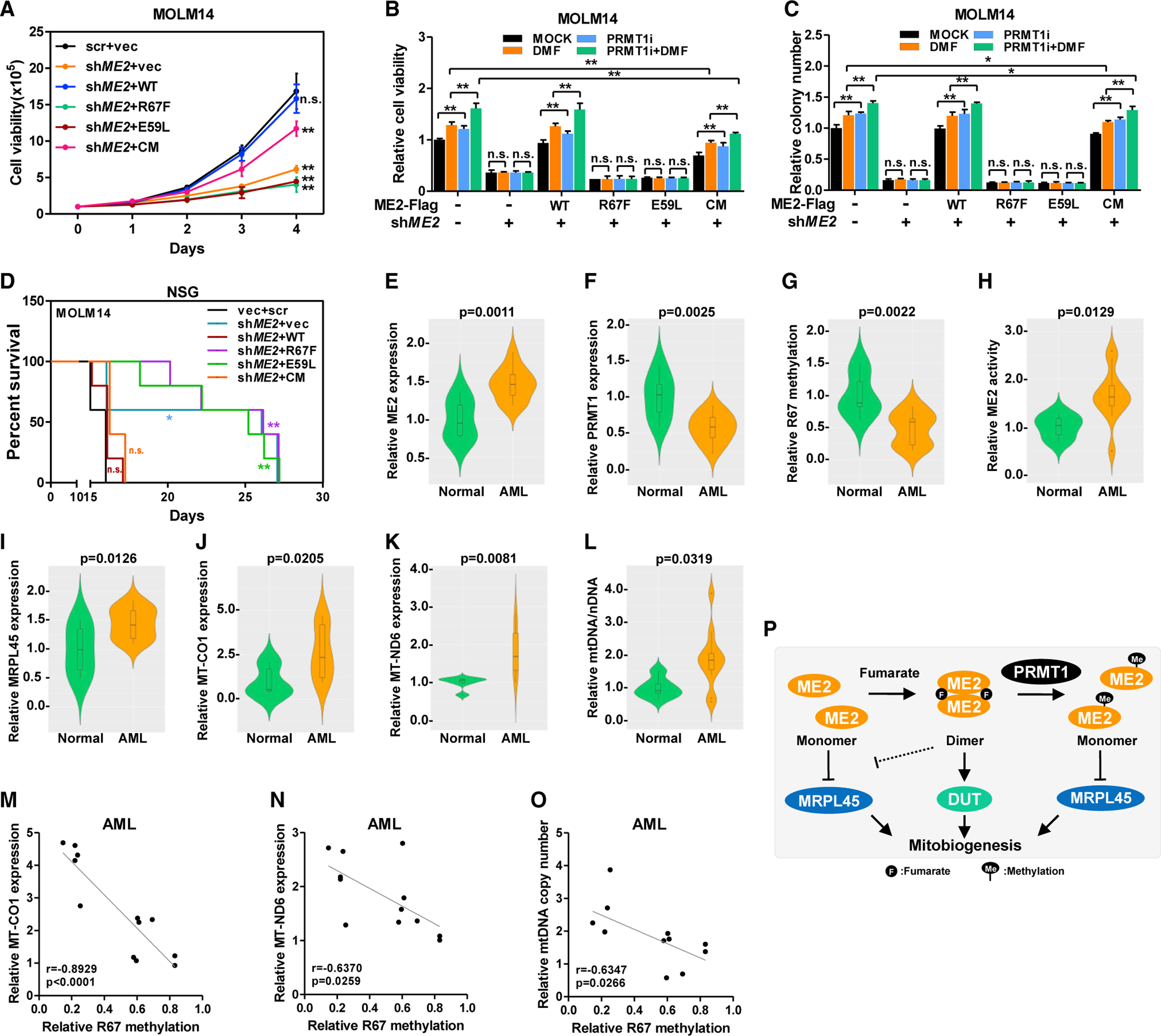
Mitochondria have an independent genome (mtDNA) and protein synthesis machinery that coordinately activate for mitochondrial generation. Here, we report that the Krebs cycle intermediate fumarate links metabolism to mitobiogenesis through binding to malic enzyme 2 (ME2). Mechanistically, fumarate binds ME2 with two complementary consequences. First, promoting the formation of ME2 dimers, which activate deoxyuridine 5'-triphosphate nucleotidohydrolase (DUT). DUT fosters thymidine generation and an increase of mtDNA. Second, fumarate-induced ME2 dimers abrogate ME2 monomer binding to mitochondrial ribosome protein L45, freeing it for mitoribosome assembly and mtDNA-encoded protein production. Methylation of the ME2-fumarate binding site by protein arginine methyltransferase-1 inhibits fumarate signaling to constrain mitobiogenesis. Notably, acute myeloid leukemia is highly dependent on mitochondrial function and is sensitive to targeting of the fumarate-ME2 axis. Therefore, mitobiogenesis can be manipulated in normal and malignant cells through ME2, an unanticipated governor of mitochondrial biomass production that senses nutrient availability through fumarate.
Keywords: acute myeloid leukemia; arginine methylation; deoxyuridine 5′-triphosphate nucleotidohydrolase; fumarate; malic enzyme 2; mitobiogenesis; mitochondrial ribosome; mitochondrial ribosome protein L45; protein arginine methyltransferase 1.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.T.S. declares the following competing interests: Novartis, sponsored research; Agios, director and equity holder; Editas Medicines, director and equity holder; Fate Therapeutics, founder and equity holder; Magenta Therapeutics, director, founder, equity holder, and consultant; Clear Creek Bio, director, equity holder, and consultant; LifeVaultBio, director and equity holder; Fog Pharma, consultant; VCanbio, consultant.
Figures
References
-
- Baccelli I, Gareau Y, Lehnertz B, Gingras S, Spinella JF, Corneau S, Mayotte N, Girard S, Frechette M, Blouin-Chagnon V, et al. (2019). Mubritinib targets the electron transport chain complex I and reveals the landscape of OXPHOS dependency in acute myeloid leukemia. Cancer Cell 36, 84–99.e8. - PubMed
-
- Bardella C, Olivero M, Lorenzato A, Geuna M, Adam J, O’Flaherty L, Rustin P, Tomlinson I, Pollard PJ, and Di Renzo MF (2012). Cells lacking the fumarase tumor suppressor are protected from apoptosis through a hypoxia-inducible factor-independent, AMPK-dependent mechanism. Mol. Cell. Biol. 32, 3081–3094. - PMC - PubMed
-
- Blanc RS, and Richard S (2017). Arginine methylation: the coming of age. Mol. Cell 65, 8–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
