

Oxidation of trimethylamine to trimethylamine N-oxide facilitates high hydrostatic pressure tolerance in a generalist bacterial lineage
- PMID: 33771875
- PMCID: PMC7997507
- DOI: 10.1126/sciadv.abf9941
Oxidation of trimethylamine to trimethylamine N-oxide facilitates high hydrostatic pressure tolerance in a generalist bacterial lineage
Abstract
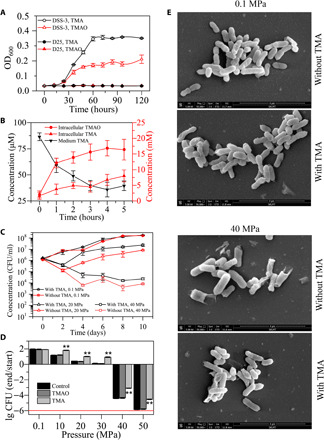
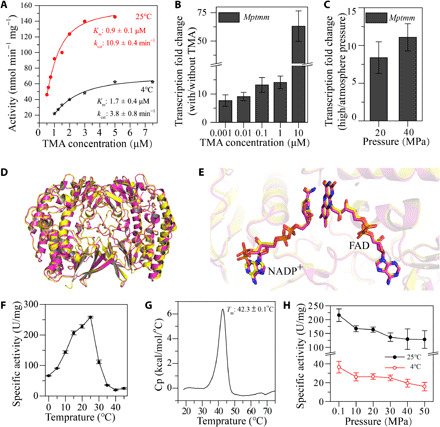
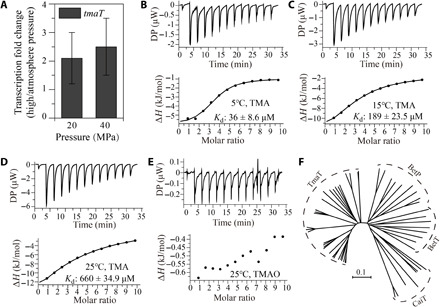
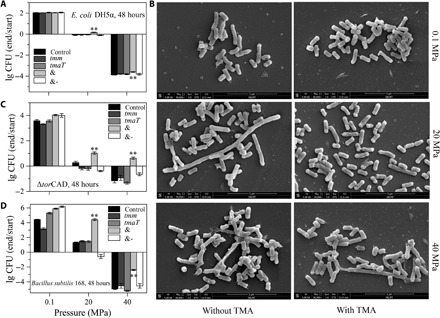
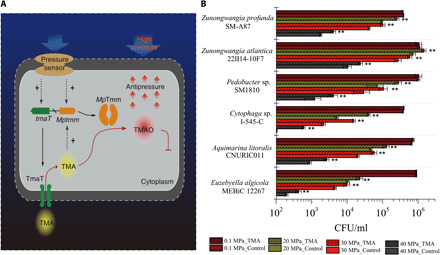
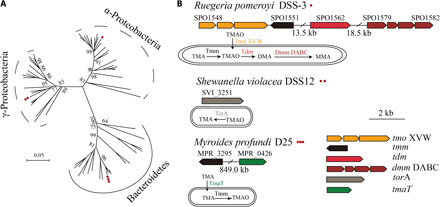
High hydrostatic pressure (HHP) is a characteristic environmental factor of the deep ocean. However, it remains unclear how piezotolerant bacteria adapt to HHP. Here, we identify a two-step metabolic pathway to cope with HHP stress in a piezotolerant bacterium. Myroides profundi D25T, obtained from a deep-sea sediment, can take up trimethylamine (TMA) through a previously unidentified TMA transporter, TmaT, and oxidize intracellular TMA into trimethylamine N-oxide (TMAO) by a TMA monooxygenase, MpTmm. The produced TMAO is accumulated in the cell, functioning as a piezolyte, improving both growth and survival at HHP. The function of the TmaT-MpTmm pathway was further confirmed by introducing it into Escherichia coli and Bacillus subtilis Encoded TmaT-like and MpTmm-like sequences extensively exist in marine metagenomes, and other marine Bacteroidetes bacteria containing genes encoding TmaT-like and MpTmm-like proteins also have improved HHP tolerance in the presence of TMA, implying the universality of this HHP tolerance strategy in marine Bacteroidetes.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Lundsten L., Schlining K. L., Frasier K., Johnson S. B., Kuhnz L. A., Harvey J. B. J., Clague G., Vrijenhoek R. C., Time-series analysis of six whale-fall communities in Monterey Canyon, California, USA. Deep Sea Res. Part I Oceanogr. Res. Papers 57, 1573–1584 (2010).
-
- Newell C. R., Pilskaln C. H., Robinson S. M., MacDonald B. A., The contribution of marine snow to the particle food supply of the benthic suspension feeder, Mytilus edulis. J. Exp. Mar. Biol. Ecol. 321, 109–124 (2005).
-
- Oger P. M., Jebbar M., The many ways of coping with pressure. Res. Microbiol. 161, 799–809 (2010). - PubMed
-
- Jebbar M., Franzetti B., Girard E., Oger P., Microbial diversity and adaptation to high hydrostatic pressure in deep-sea hydrothermal vents prokaryotes. Extremophiles 19, 721–740 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
