

Proteasome stress in skeletal muscle mounts a long-range protective response that delays retinal and brain aging
- PMID: 33773104
- PMCID: PMC8172468
- DOI: 10.1016/j.cmet.2021.03.005
Proteasome stress in skeletal muscle mounts a long-range protective response that delays retinal and brain aging
Abstract
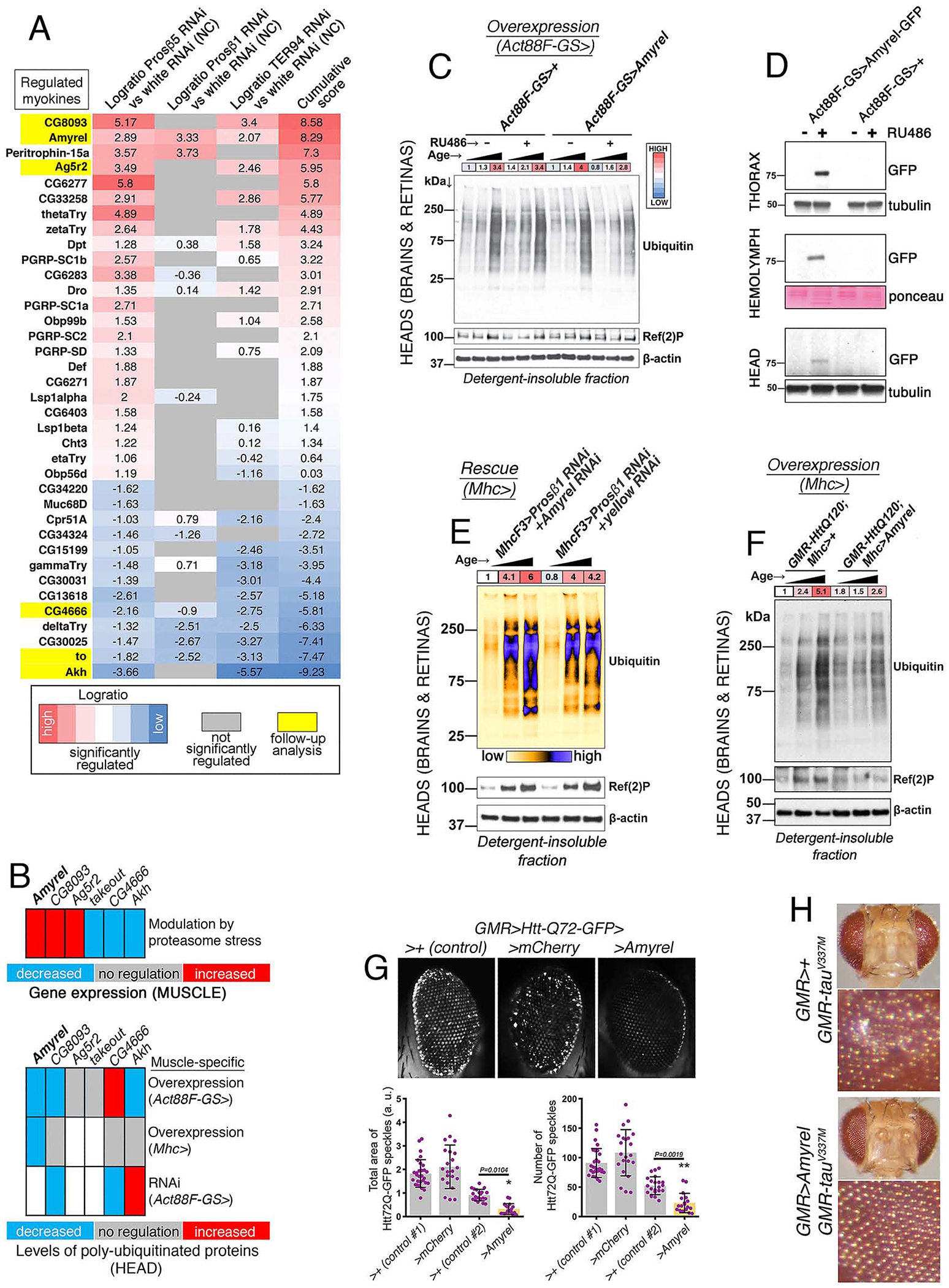
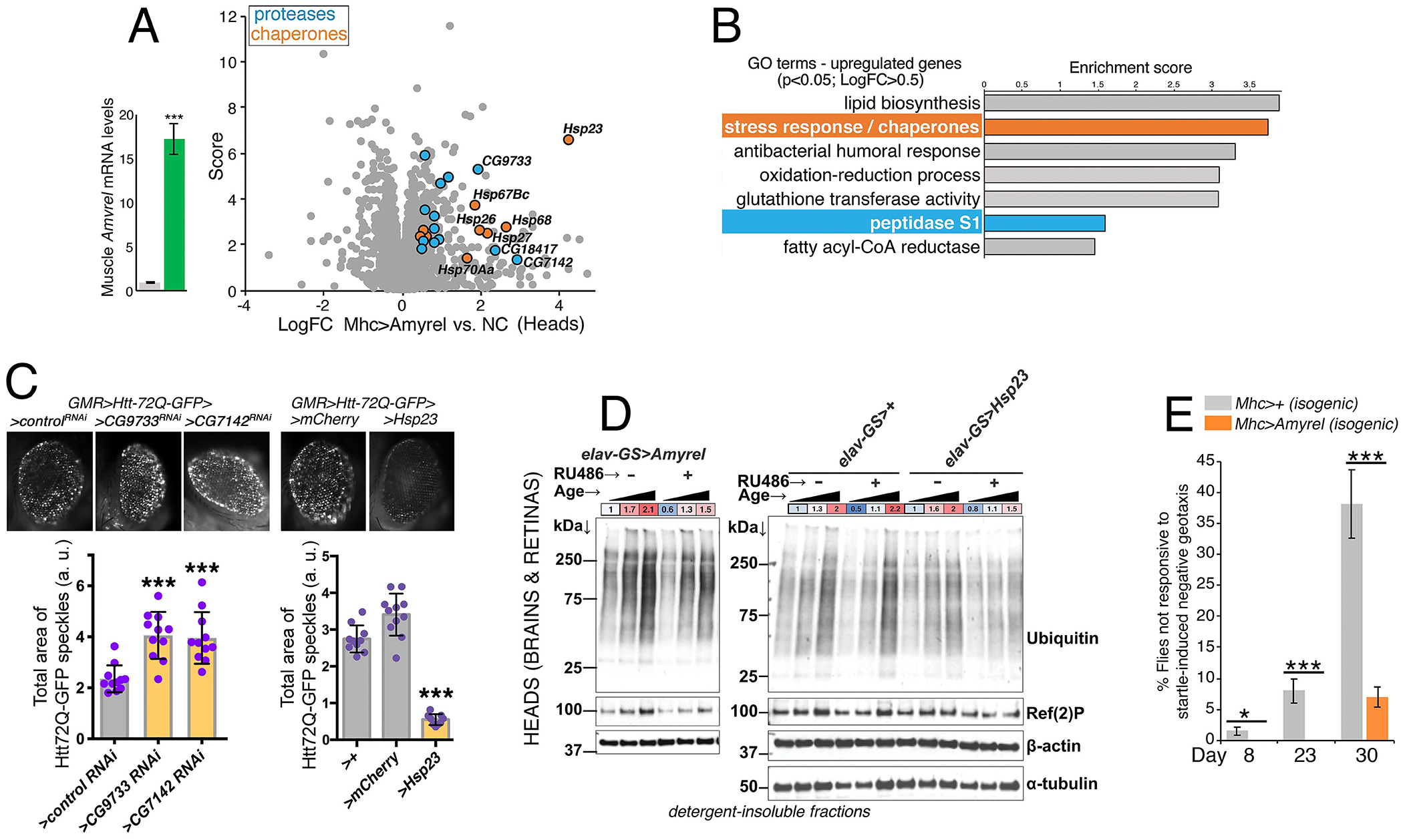
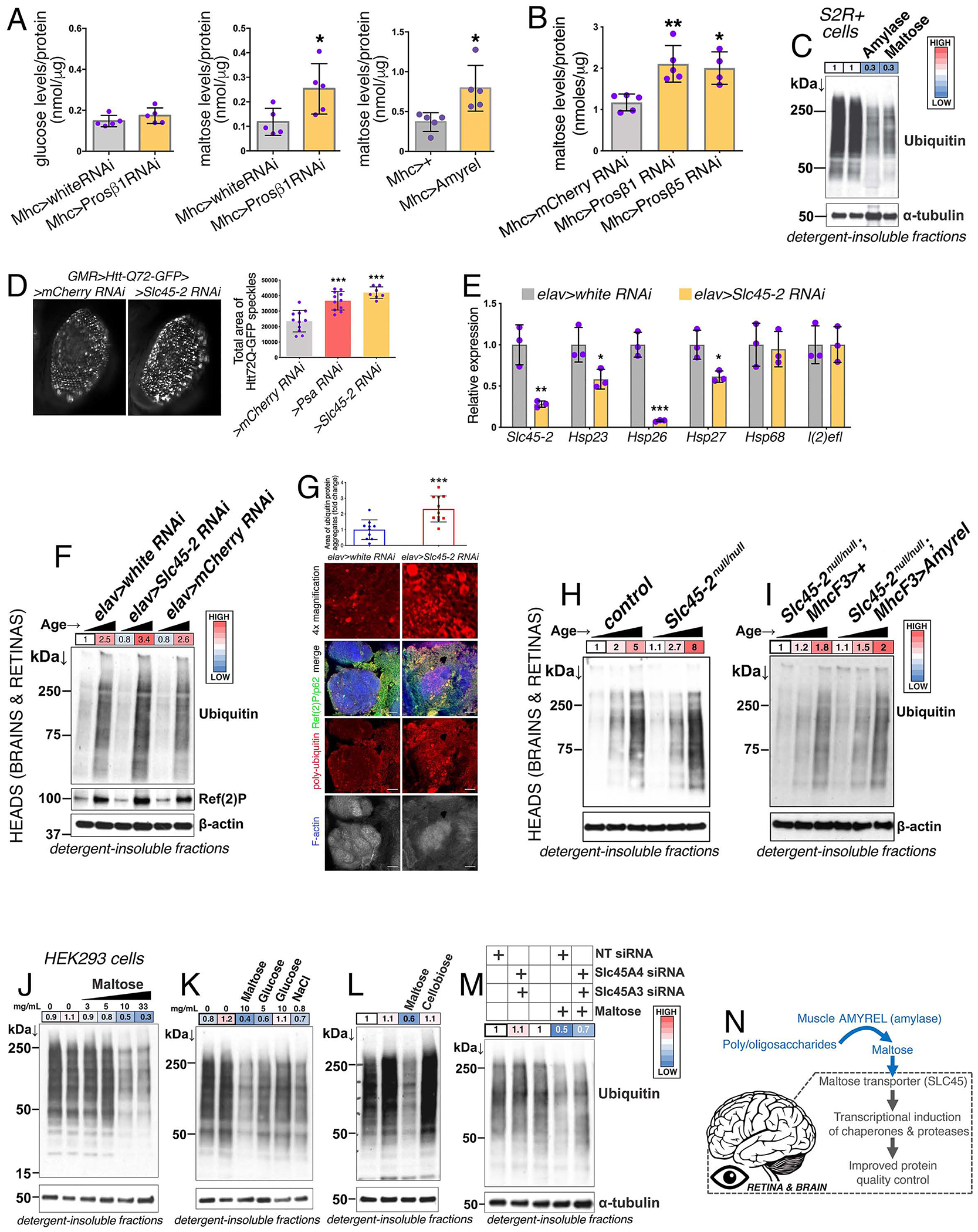
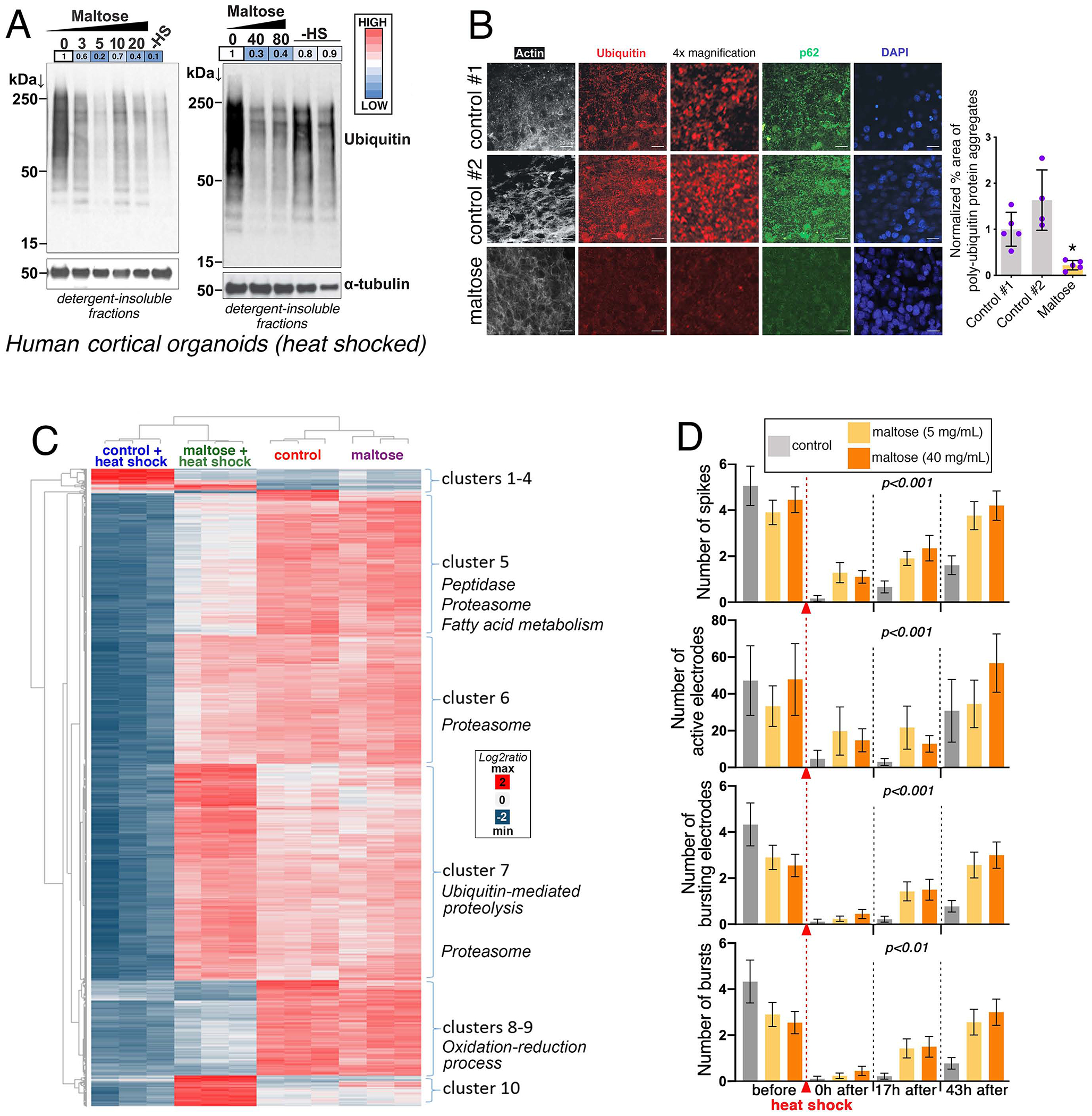
Neurodegeneration in the central nervous system (CNS) is a defining feature of organismal aging that is influenced by peripheral tissues. Clinical observations indicate that skeletal muscle influences CNS aging, but the underlying muscle-to-brain signaling remains unexplored. In Drosophila, we find that moderate perturbation of the proteasome in skeletal muscle induces compensatory preservation of CNS proteostasis during aging. Such long-range stress signaling depends on muscle-secreted Amyrel amylase. Mimicking stress-induced Amyrel upregulation in muscle reduces age-related accumulation of poly-ubiquitinated proteins in the brain and retina via chaperones. Preservation of proteostasis stems from the disaccharide maltose, which is produced via Amyrel amylase activity. Correspondingly, RNAi for SLC45 maltose transporters reduces expression of Amyrel-induced chaperones and worsens brain proteostasis during aging. Moreover, maltose preserves proteostasis and neuronal activity in human brain organoids challenged by thermal stress. Thus, proteasome stress in skeletal muscle hinders retinal and brain aging by mounting an adaptive response via amylase/maltose.
Keywords: aging; amylase; brain organoids; maltose; muscle-to-brain signaling; muscle-to-retina signaling; myokine; proteasome; proteostasis; stress response.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.R. and F.D. are named co-inventors of a pending U.S. provisional patent application based in part on the research reported in this paper.
Figures



Comment in
-
Brains and brawn: Stress-induced myokine abates nervous system aging.Cell Metab. 2021 Jun 1;33(6):1067-1069. doi: 10.1016/j.cmet.2021.05.007. Cell Metab. 2021. PMID: 34077712
References
-
- Agudelo LZ, Femenia T, Orhan F, Porsmyr-Palmertz M, Goiny M, Martinez-Redondo V, Correia JC, Izadi M, Bhat M, Schuppe-Koistinen I, et al. (2014). Skeletal muscle PGC-1alpha1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 159, 33–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
