

Principles of Alternating Access in Multidrug and Toxin Extrusion (MATE) Transporters
- PMID: 33774036
- PMCID: PMC8292195
- DOI: 10.1016/j.jmb.2021.166959
Principles of Alternating Access in Multidrug and Toxin Extrusion (MATE) Transporters
Abstract
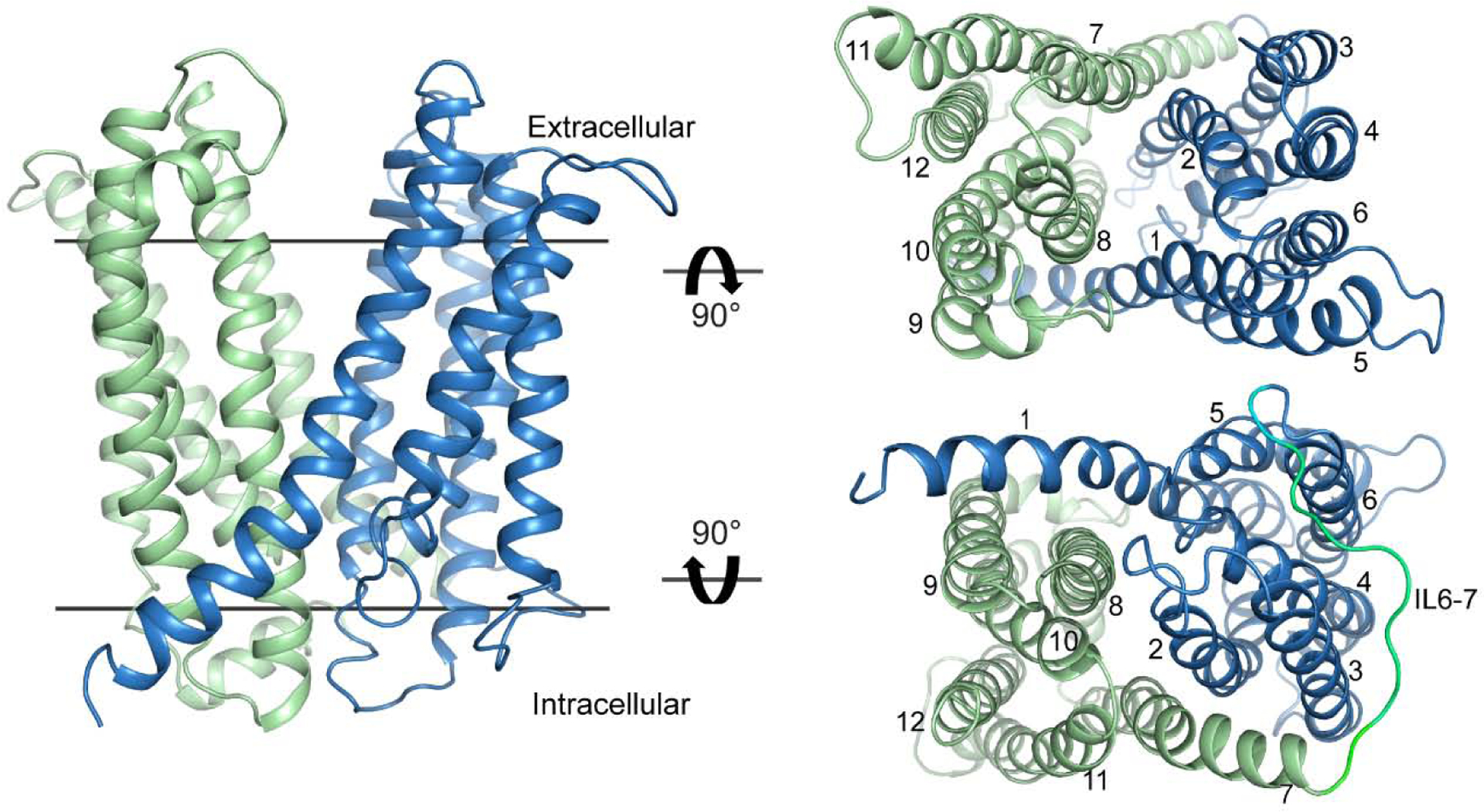
The multidrug and toxin extrusion (MATE) transporters catalyze active efflux of a broad range of chemically- and structurally-diverse compounds including antimicrobials and chemotherapeutics, thus contributing to multidrug resistance in pathogenic bacteria and cancers. Multiple methodological approaches have been taken to investigate the structural basis of energy transduction and substrate translocation in MATE transporters. Crystal structures representing members from all three MATE subfamilies have been interpreted within the context of an alternating access mechanism that postulates occupation of distinct structural intermediates in a conformational cycle powered by electrochemical ion gradients. Here we review the structural biology of MATE transporters, integrating the crystallographic models with biophysical and computational studies to define the molecular determinants that shape the transport energy landscape. This holistic analysis highlights both shared and disparate structural and functional features within the MATE family, which underpin an emerging theme of mechanistic diversity within the framework of a conserved structural scaffold.
Keywords: DinF; MATE; NorM; PfMATE; alternating access; antiport; multidrug resistance.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures










References
-
- Du D, van Veen HW, Murakami S, Pos KM & Luisi BF (2015). Structure, mechanism and cooperation of bacterial multidrug transporters. Curr Opin Struct Biol 33, 76–91. - PubMed
-
- Higgins CF (2007). Multiple molecular mechanisms for multidrug resistance transporters. Nature 446, 749–57. - PubMed
-
- Alibert S, N’Gompaza Diarra J, Hernandez J, Stutzmann A, Fouad M, Boyer G & Pages JM (2017). Multidrug efflux pumps and their role in antibiotic and antiseptic resistance: a pharmacodynamic perspective. Expert Opin Drug Metab Toxicol 13, 301–309. - PubMed
-
- Gottesman MM, Fojo T & Bates SE (2002). Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer 2, 48–58. - PubMed
-
- Blair JM, Webber MA, Baylay AJ, Ogbolu DO & Piddock LJ (2014). Molecular mechanisms of antibiotic resistance. Nat Rev Microbiol 13, 42–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
