

Rqc1 and other yeast proteins containing highly positively charged sequences are not targets of the RQC complex
- PMID: 33774050
- PMCID: PMC8102910
- DOI: 10.1016/j.jbc.2021.100586
Rqc1 and other yeast proteins containing highly positively charged sequences are not targets of the RQC complex
Abstract
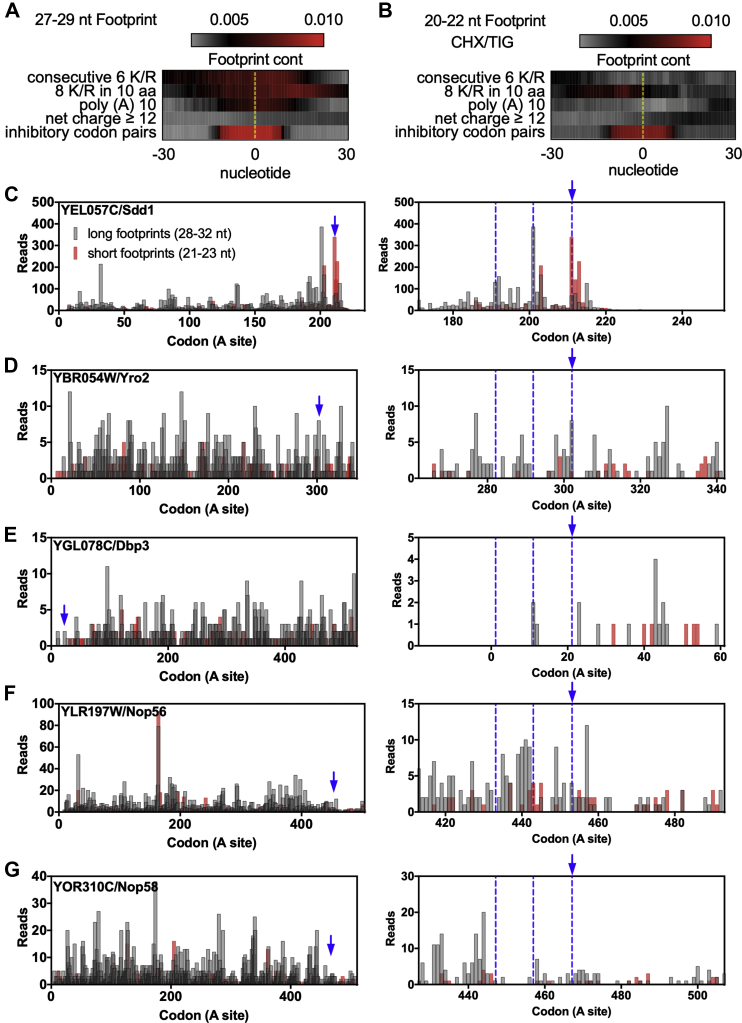
Previous work has suggested that highly positively charged protein segments coded by rare codons or poly (A) stretches induce ribosome stalling and translational arrest through electrostatic interactions with the negatively charged ribosome exit tunnel, leading to inefficient elongation. This arrest leads to the activation of the Ribosome Quality Control (RQC) pathway and results in low expression of these reporter proteins. However, the only endogenous yeast proteins known to activate the RQC are Rqc1, a protein essential for RQC function, and Sdd1, a protein with unknown function, both of which contain polybasic sequences. To explore the generality of this phenomenon, we investigated whether the RQC complex controls the expression of other proteins with polybasic sequences. We showed by ribosome profiling data analysis and western blot that proteins containing polybasic sequences similar to, or even more positively charged than those of Rqc1 and Sdd1, were not targeted by the RQC complex. We also observed that the previously reported Ltn1-dependent regulation of Rqc1 is posttranslational, independent of the RQC activity. Taken together, our results suggest that RQC should not be regarded as a general regulatory pathway for the expression of highly positively charged proteins in yeast.
Keywords: poly (A) tracts; polybasic sequences; protein synthesis; ribosome; ribosome profiling; ribosome quality control; stalled polypeptides; translation; translation control; ubiquitin ligase.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures




References
-
- Lu J., Kobertz W.R., Deutsch C. Mapping the electrostatic potential within the ribosomal exit tunnel. J. Mol. Biol. 2007;371:1378–1391. - PubMed
-
- Frischmeyer P.A., van Hoof A., O’Donnell K., Guerrerio A.L., Parker R., Dietz H.C. An mRNA surveillance mechanism that eliminates transcripts lacking termination codons. Science. 2002;295:2258–2261. - PubMed
-
- van Hoof A., Frischmeyer P.A., Dietz H.C., Parker R. Exosome-mediated recognition and degradation of mRNAs lacking a termination codon. Science. 2002;295:2262–2264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
