

Heart Enhancers: Development and Disease Control at a Distance
- PMID: 33777110
- PMCID: PMC7987942
- DOI: 10.3389/fgene.2021.642975
Heart Enhancers: Development and Disease Control at a Distance
Abstract
Bound by lineage-determining transcription factors and signaling effectors, enhancers play essential roles in controlling spatiotemporal gene expression profiles during development, homeostasis and disease. Recent synergistic advances in functional genomic technologies, combined with the developmental biology toolbox, have resulted in unprecedented genome-wide annotation of heart enhancers and their target genes. Starting with early studies of vertebrate heart enhancers and ending with state-of-the-art genome-wide enhancer discovery and testing, we will review how studying heart enhancers in metazoan species has helped inform our understanding of cardiac development and disease.
Keywords: cardiac gene expression; comparative genomics; enhancer; epigenomics and epigenetics; gene regulation; transcription factor (TF).
Copyright © 2021 Yuan, Scott and Wilson.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
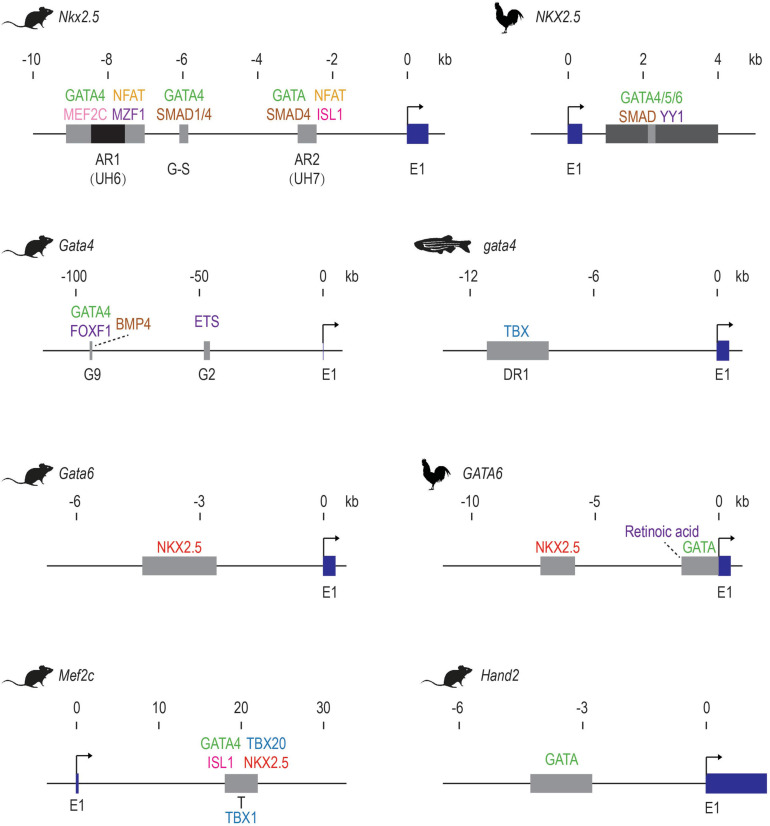
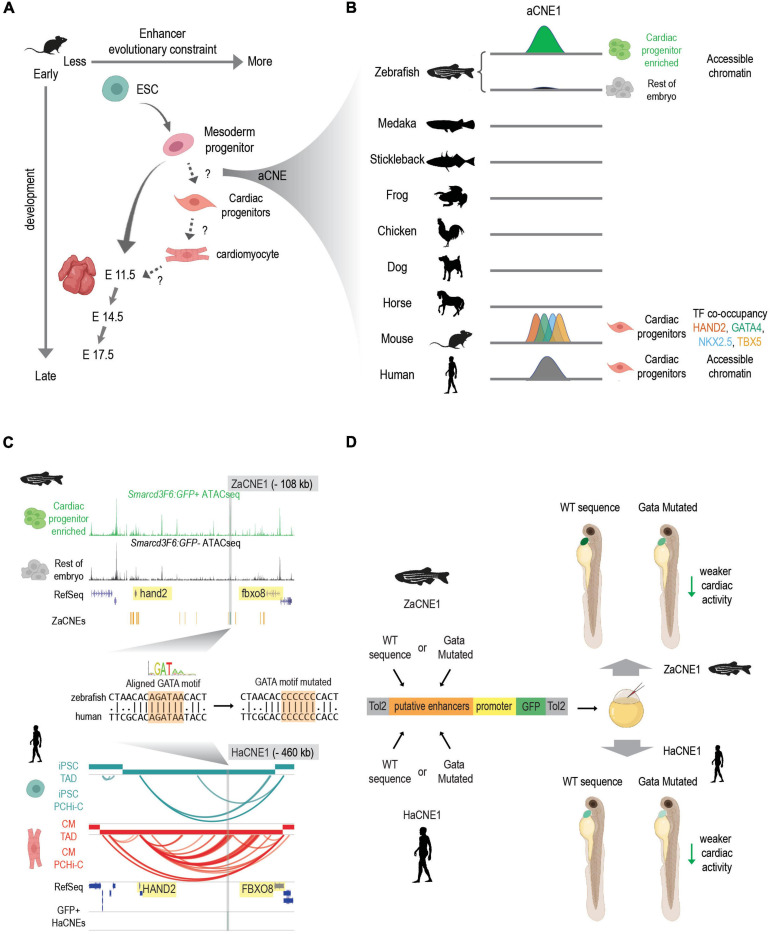
References
-
- Ahmad S. M., Busser B. W., Huang D., Cozart E. J., Michaud S., Zhu X., et al. (2014). Machine learning classification of cell-specific cardiac enhancers uncovers developmental subnetworks regulating progenitor cell division and cell fate specification. Dev. 141 878–888. 10.1242/dev.101709 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous