

Regulation of Superoxide by BAP31 through Its Effect on p22phox and Keap1/Nrf2/HO-1 Signaling Pathway in Microglia
- PMID: 33777312
- PMCID: PMC7969104
- DOI: 10.1155/2021/1457089
Regulation of Superoxide by BAP31 through Its Effect on p22phox and Keap1/Nrf2/HO-1 Signaling Pathway in Microglia
Abstract
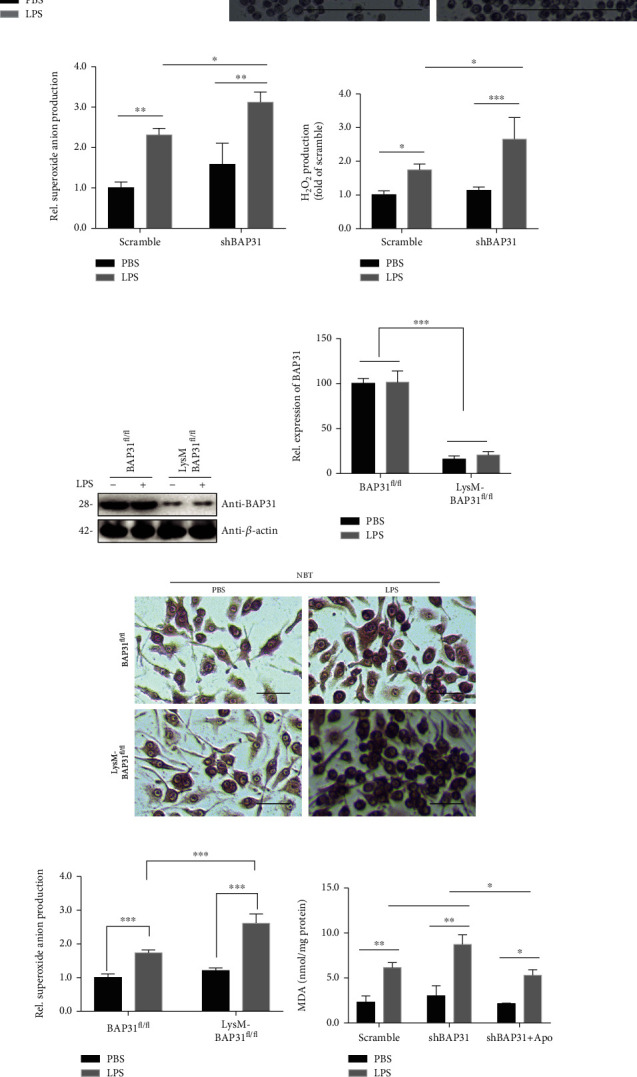
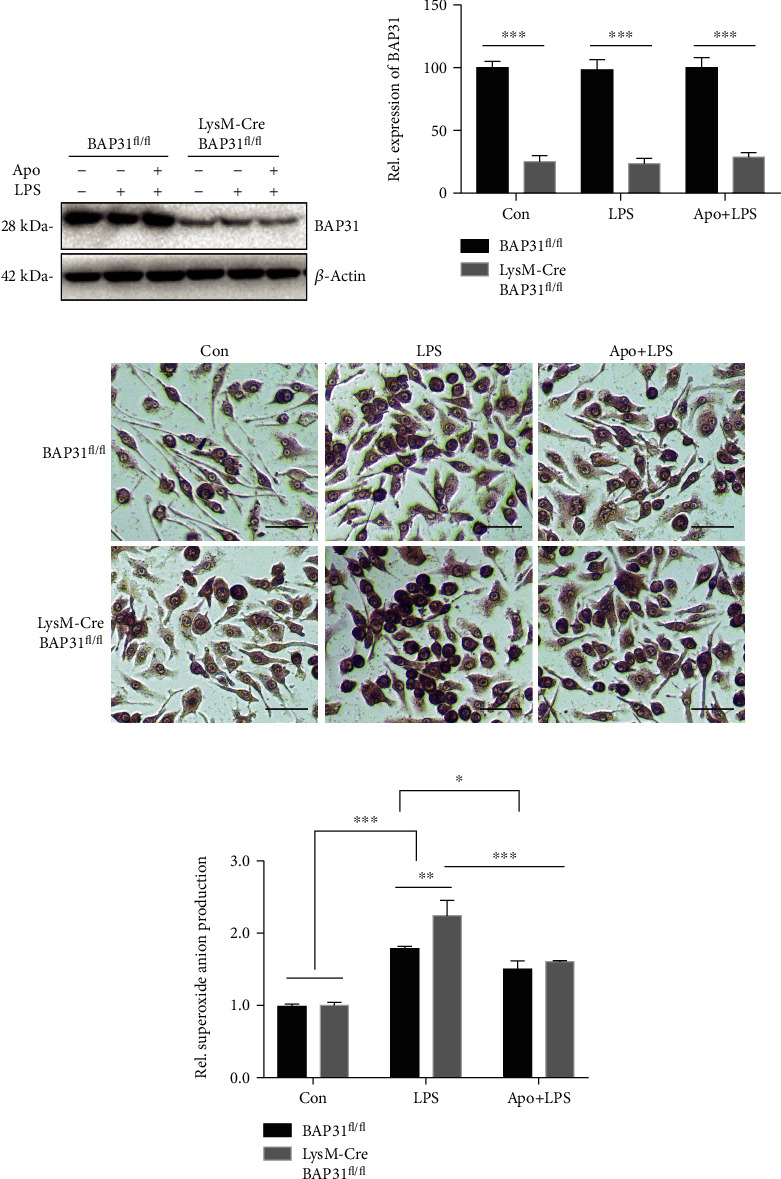
Reactive oxygen species (ROS) production by activation of microglia is considered to be a major cause of neuronal dysfunction, which can lead to damage and death through direct oxidative damage to neuronal macromolecules or derangement of neuronal redox signaling circuits. BAP31, an integral ER membrane protein, has been defined as a regulatory molecule in the CNS. Our latest studies have found that BAP31 deficiency leads to activation of microglia. In this study, we discovered that BAP31 deficiency upregulated LPS-induced superoxide anion production in BV2 cells and mice by upregulating the expression level of p22phox and by inhibiting the activation of Nrf2-HO-1 signaling. Knockdown of p22phox/keap1 or use of an NADPH oxidase inhibitor (apocynin) reversed the production of superoxide anion and inflammatory cytokines, which then reduced neuronal damage and death in vitro and in vivo. These results suggest that BAP31 deficiency contributes to microglia-related superoxide anion production and neuroinflammation through p22phox and keap1. Furthermore, the excess superoxide anion cooperated with inflammatory cytokines to induce the damage and death of neurons. Thus, we determined that BAP31 is an important regulator in superoxide anion production and neuroinflammation, and the downstream regulators or agonists of BAP31 could therefore be considered as potential therapeutic targets in microglial-related superoxide anion production and neuroinflammation.
Copyright © 2021 Xia Liu et al.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures









Similar articles
-
A Novel Interaction of Translocator Protein 18 kDa (TSPO) with NADPH Oxidase in Microglia.Mol Neurobiol. 2020 Nov;57(11):4467-4487. doi: 10.1007/s12035-020-02042-w. Epub 2020 Aug 2. Mol Neurobiol. 2020. PMID: 32743737 Free PMC article.
-
Andrographolide Activates Keap1/Nrf2/ARE/HO-1 Pathway in HT22 Cells and Suppresses Microglial Activation by Aβ42 through Nrf2-Related Inflammatory Response.Mediators Inflamm. 2017;2017:5906189. doi: 10.1155/2017/5906189. Epub 2017 Mar 8. Mediators Inflamm. 2017. PMID: 28373747 Free PMC article.
-
BAP31 regulates IRAK1-dependent neuroinflammation in microglia.J Neuroinflammation. 2019 Dec 28;16(1):281. doi: 10.1186/s12974-019-1661-7. J Neuroinflammation. 2019. PMID: 31883536 Free PMC article.
-
NOX inhibitors as a therapeutic strategy for stroke and neurodegenerative disease.Curr Drug Targets. 2012 Feb;13(2):199-206. doi: 10.2174/138945012799201676. Curr Drug Targets. 2012. PMID: 22204319 Review.
-
Microglia antioxidant systems and redox signalling.Br J Pharmacol. 2017 Jun;174(12):1719-1732. doi: 10.1111/bph.13426. Epub 2016 Mar 3. Br J Pharmacol. 2017. PMID: 26754582 Free PMC article. Review.
Cited by
-
Interplay of α-Synuclein Oligomers and Endoplasmic Reticulum Stress in Parkinson'S Disease: Insights into Cellular Dysfunctions.Inflammation. 2025 Aug;48(4):1590-1606. doi: 10.1007/s10753-024-02156-6. Epub 2024 Oct 9. Inflammation. 2025. PMID: 39382817 Review.
-
HNF4A-BAP31-VDAC1 axis synchronously regulates cell proliferation and ferroptosis in gastric cancer.Cell Death Dis. 2023 Jun 9;14(6):356. doi: 10.1038/s41419-023-05868-z. Cell Death Dis. 2023. PMID: 37296105 Free PMC article.
-
The Effects of Antioxidant Approved Drugs and Under Investigation Compounds with Potential of Improving Sleep Disorders and their Associated Comorbidities Associated with Oxidative Stress and Inflammation.Mini Rev Med Chem. 2025;25(10):795-815. doi: 10.2174/0113895575360959250117073046. Mini Rev Med Chem. 2025. PMID: 39957704 Review.
References
-
- Han B. H., Zhou M. L., Johnson A. W., et al. Contribution of reactive oxygen species to cerebral amyloid angiopathy, vasomotor dysfunction, and microhemorrhage in aged Tg2576 mice. Proceedings of the National Academy of Sciences of the United States of America. 2015;112(8):E881–E890. doi: 10.1073/pnas.1414930112. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
