

The Role of Dopamine in Associative Learning in Drosophila: An Updated Unified Model
- PMID: 33779893
- PMCID: PMC8192648
- DOI: 10.1007/s12264-021-00665-0
The Role of Dopamine in Associative Learning in Drosophila: An Updated Unified Model
Abstract
Learning to associate a positive or negative experience with an unrelated cue after the presentation of a reward or a punishment defines associative learning. The ability to form associative memories has been reported in animal species as complex as humans and as simple as insects and sea slugs. Associative memory has even been reported in tardigrades [1], species that diverged from other animal phyla 500 million years ago. Understanding the mechanisms of memory formation is a fundamental goal of neuroscience research. In this article, we work on resolving the current contradictions between different Drosophila associative memory circuit models and propose an updated version of the circuit model that predicts known memory behaviors that current models do not. Finally, we propose a model for how dopamine may function as a reward prediction error signal in Drosophila, a dopamine function that is well-established in mammals but not in insects [2, 3].
Figures


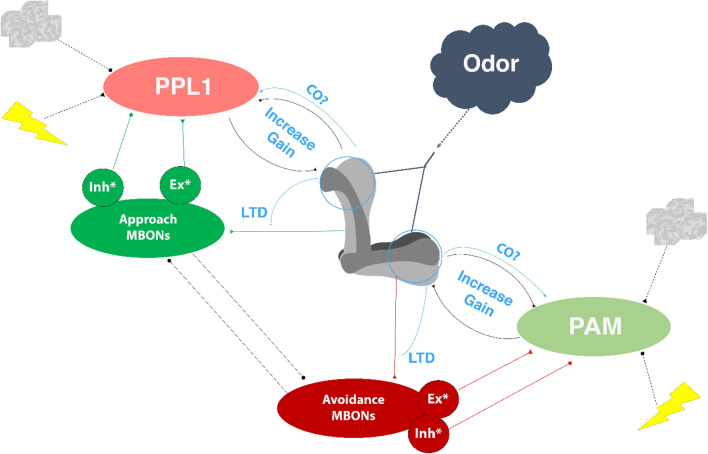



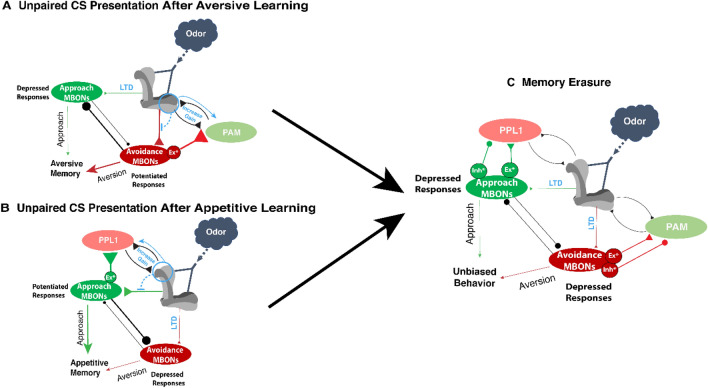



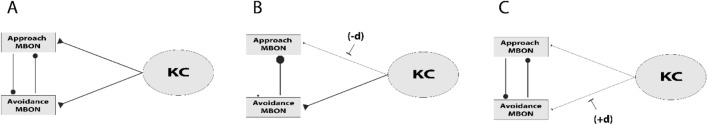

References
-
- Riemensperger T, Völler T, Stock P, Buchner E, Fiala A. Punishment prediction by dopaminergic neurons in Drosophila. Curr Biol. 2005;15:1953–1960. - PubMed
-
- Heisenberg M. Mushroom body memoir: from maps to models. Nat Rev Neurosci. 2003;4:266–275. - PubMed
-
- Keene AC, Waddell S. Drosophila olfactory memory: single genes to complex neural circuits. Nat Rev Neurosci. 2007;8:341–354. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
