

TBK1 recruitment to STING activates both IRF3 and NF-κB that mediate immune defense against tumors and viral infections
- PMID: 33785602
- PMCID: PMC8040795
- DOI: 10.1073/pnas.2100225118
TBK1 recruitment to STING activates both IRF3 and NF-κB that mediate immune defense against tumors and viral infections
Abstract
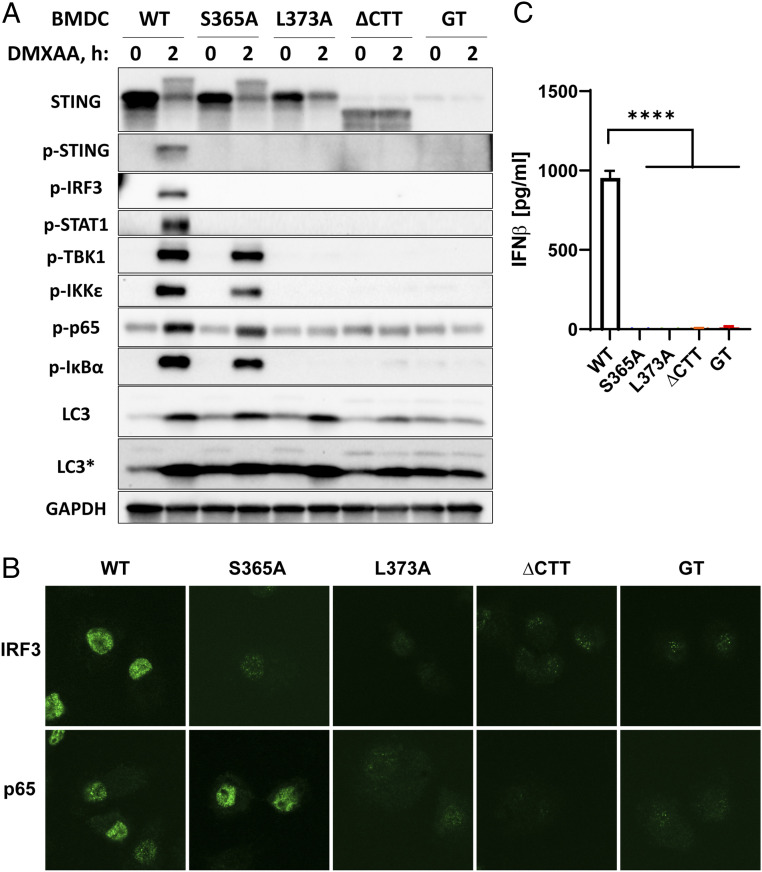
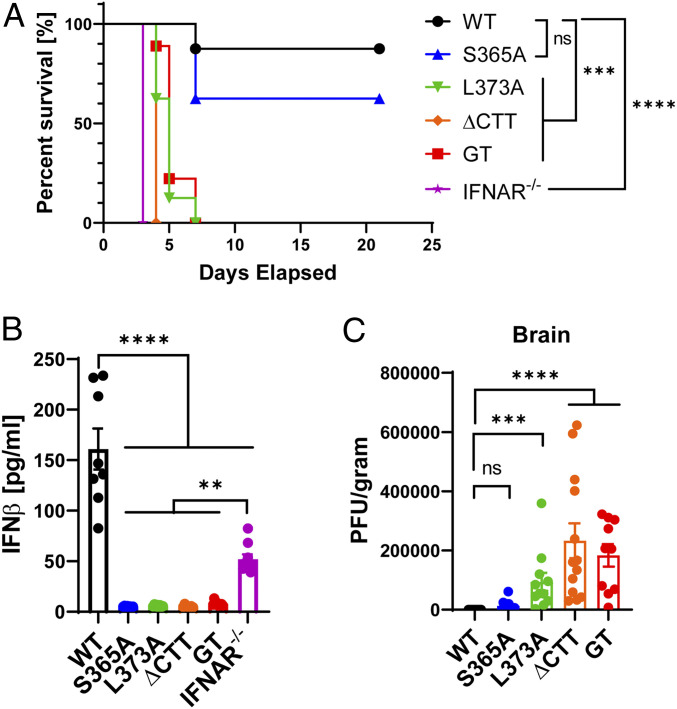
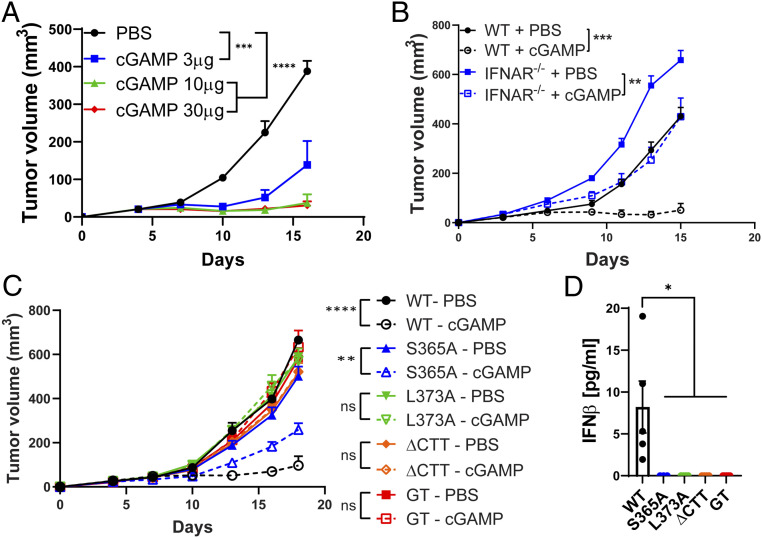
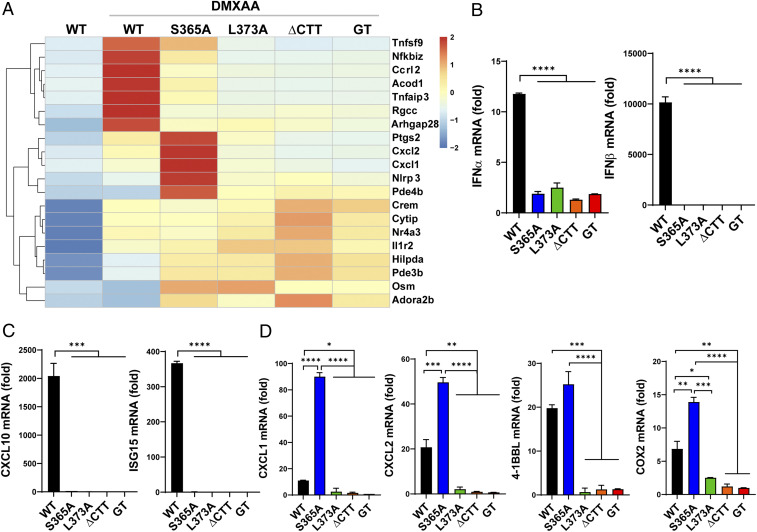
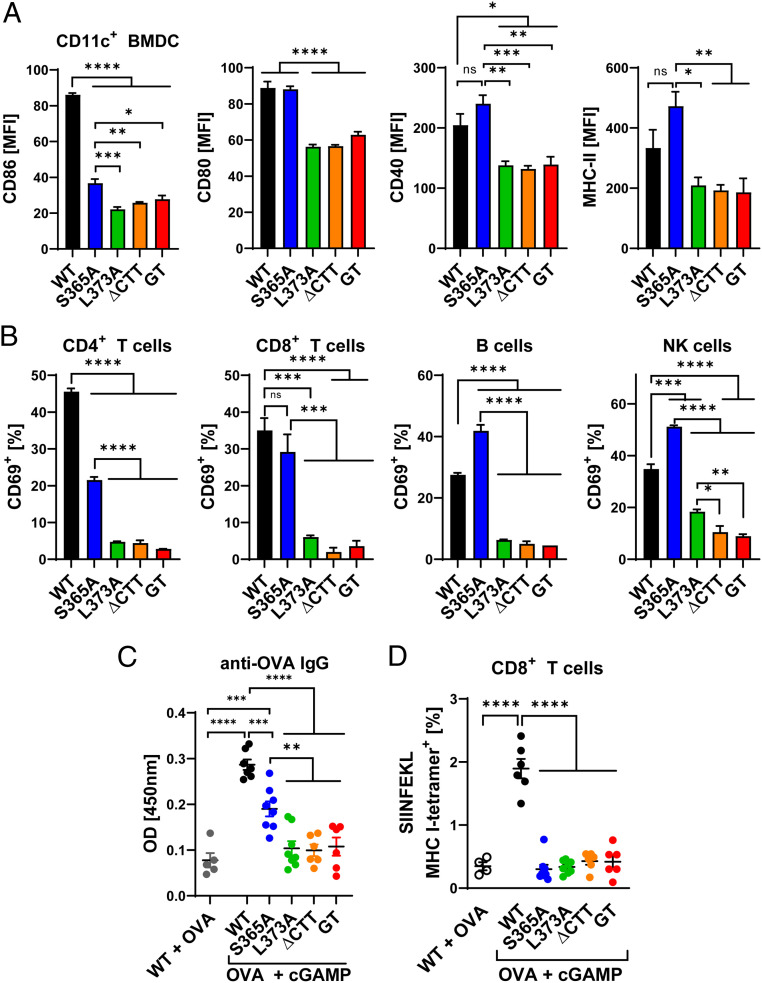
The induction of type I interferons through the transcription factor interferon regulatory factor 3 (IRF3) is considered a major outcome of stimulator of interferon genes (STING) activation that drives immune responses against DNA viruses and tumors. However, STING activation can also trigger other downstream pathways such as nuclear factor κB (NF-κB) signaling and autophagy, and the roles of interferon (IFN)-independent functions of STING in infectious diseases or cancer are not well understood. Here, we generated a STING mouse strain with a mutation (S365A) that disrupts IRF3 binding and therefore type I interferon induction but not NF-κB activation or autophagy induction. We also generated STING mice with mutations that disrupt the recruitment of TANK-binding kinase 1 (TBK1), which is important for both IRF3 and NF-κB activation but not autophagy induction (L373A or ∆CTT, which lacks the C-terminal tail). The STING-S365A mutant mice, but not L373A or ∆CTT mice, were still resistant to herpes simplex virus 1 (HSV-1) infections and mounted an antitumor response after cyclic guanosine monophosphate-adenosine monophosphate (cGAMP) treatment despite the absence of STING-induced interferons. These results demonstrate that STING can function independently of type I interferons and autophagy, and that TBK1 recruitment to STING is essential for antiviral and antitumor immunity.
Keywords: NF-κB; STING; TBK1; cGAS; interferon.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Zhong B., et al., The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 29, 538–550 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
