

Balancing Positive and Negative Selection: In Vivo Evolution of Candida lusitaniae MRR1
- PMID: 33785623
- PMCID: PMC8092287
- DOI: 10.1128/mBio.03328-20
Balancing Positive and Negative Selection: In Vivo Evolution of Candida lusitaniae MRR1
Abstract
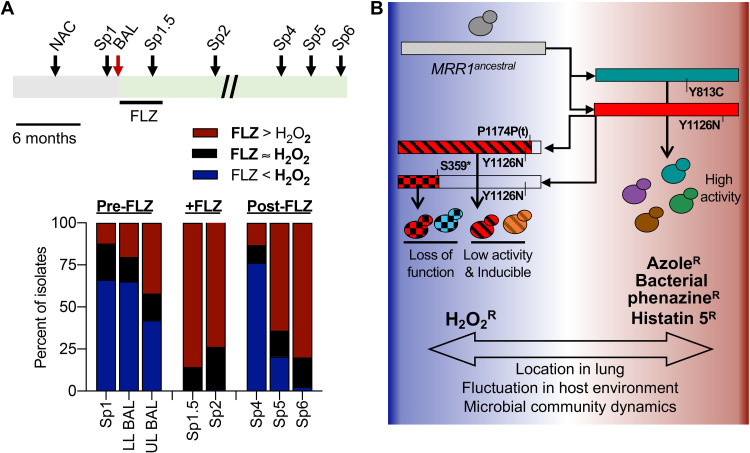
The evolution of pathogens in response to selective pressures present during chronic infections can influence their persistence and virulence and the outcomes of antimicrobial therapy. Because subpopulations within an infection can be spatially separated and the host environment can fluctuate, an appreciation of the pathways under selection may be most easily revealed through the analysis of numerous isolates from single infections. Here, we continued our analysis of a set of clonally derived Clavispora (Candida) lusitaniae isolates from a single chronic lung infection with a striking enrichment in the number of alleles of MRR1 Genetic and genomic analyses found evidence for repeated acquisition of gain-of-function mutations that conferred constitutive Mrr1 activity. In the same population, there were multiple alleles with both gain-of-function mutations and secondary suppressor mutations that either attenuated or abolished the constitutive activity, suggesting the presence of counteracting selective pressures. Our studies demonstrated trade-offs between high Mrr1 activity, which confers resistance to the antifungal fluconazole, host factors, and bacterial products through its regulation of MDR1, and resistance to hydrogen peroxide, a reactive oxygen species produced in the neutrophilic environment associated with this infection. This inverse correlation between high Mrr1 activity and hydrogen peroxide resistance was observed in multiple Candida species and in serially collected populations from this individual over 3 years. These data lead us to propose that dynamic or variable selective pressures can be reflected in population genomics and that these dynamics can complicate the drug resistance profile of the population.IMPORTANCE Understanding microbial evolution within patients is critical for managing chronic infections and understanding host-pathogen interactions. Here, our analysis of multiple MRR1 alleles in isolates from a single Clavispora (Candida) lusitaniae infection revealed the selection for both high and low Mrr1 activity. Our studies reveal trade-offs between high Mrr1 activity, which confers resistance to the commonly used antifungal fluconazole, host antimicrobial peptides, and bacterial products, and resistance to hydrogen peroxide. This work suggests that spatial or temporal differences within chronic infections can support a large amount of dynamic and parallel evolution and that Mrr1 activity is under both positive and negative selective pressure to balance different traits that are important for microbial survival.
Keywords: Candida albicans; Candida auris; Candida lusitaniae; Mrr1; chronic infection; cystic fibrosis; drug resistance; evolution; fluconazole; hydrogen peroxide; yeast.
Copyright © 2021 Demers et al.
Figures
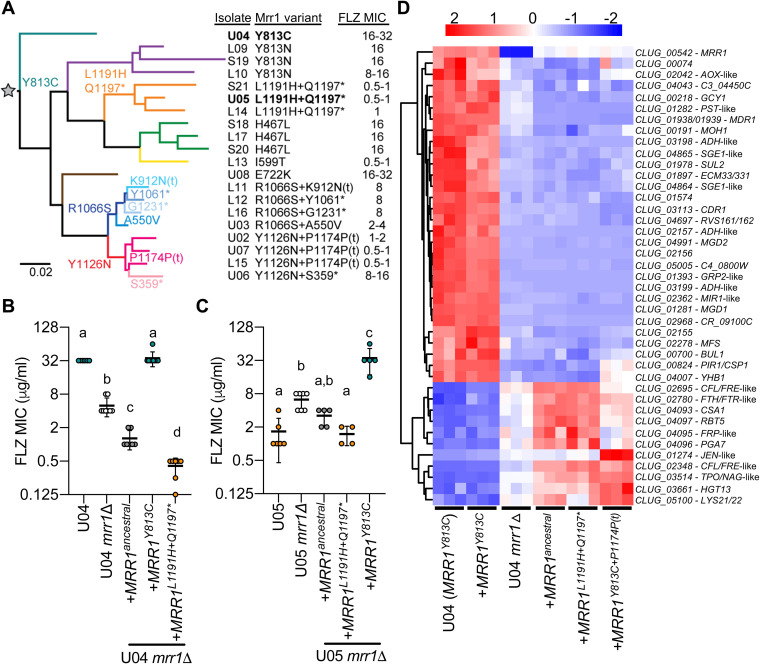
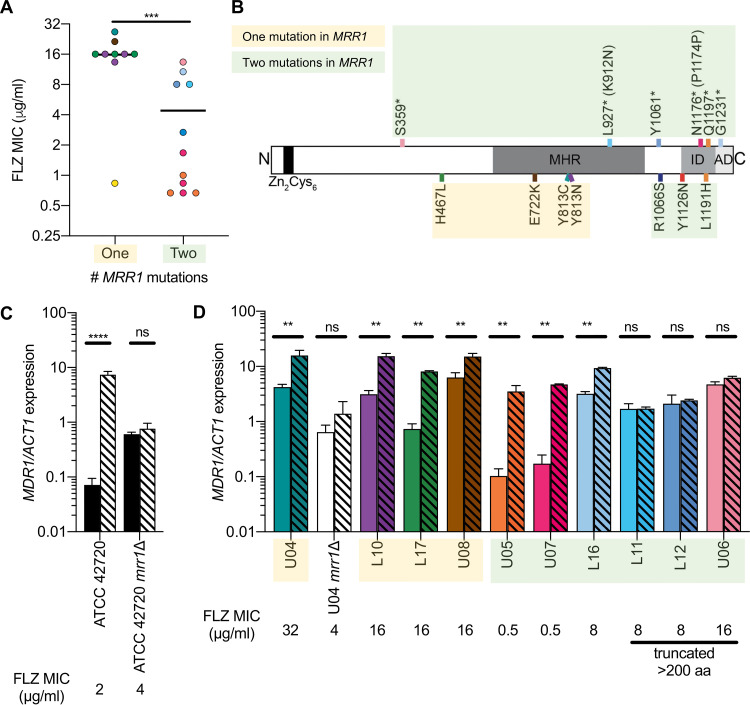
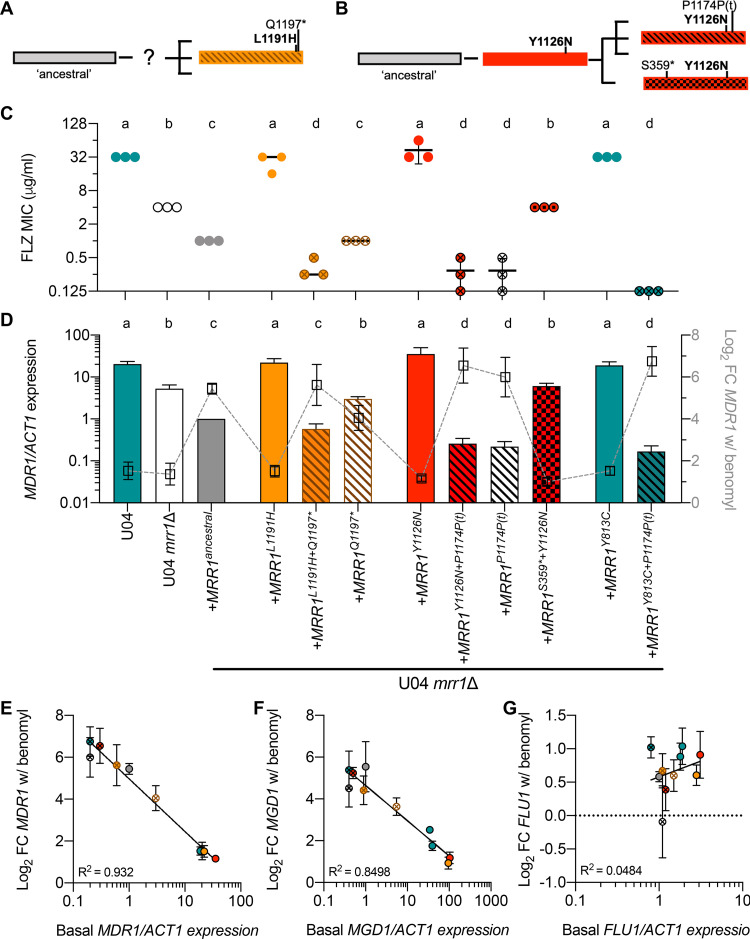
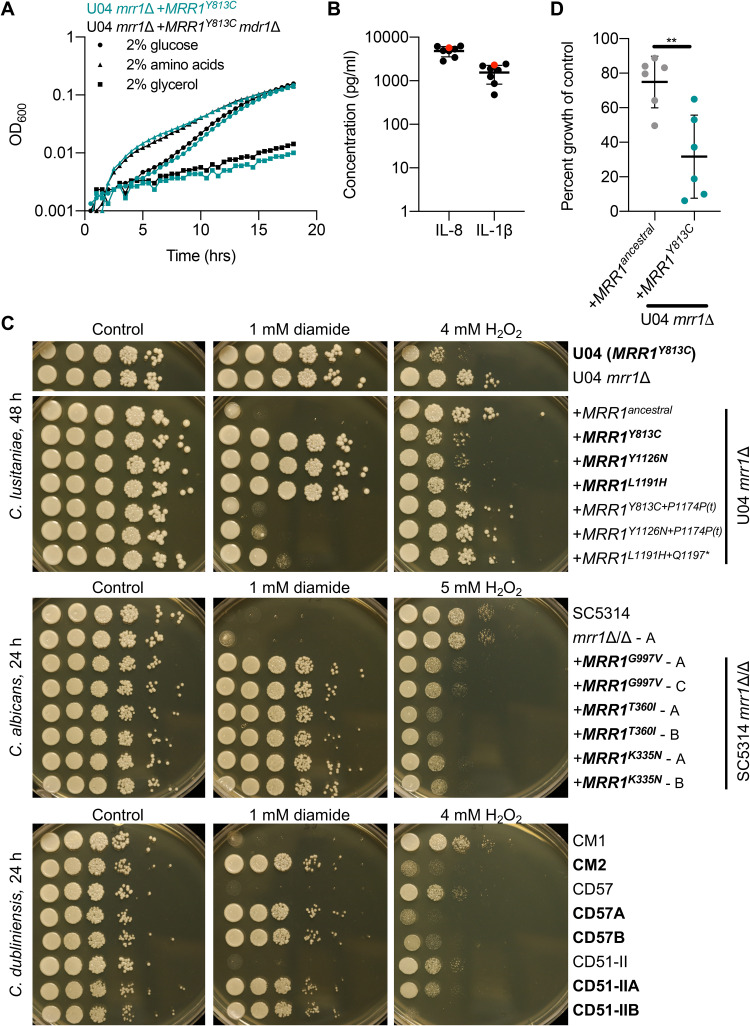
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
