

Cytosolic delivery of nucleic acids: The case of ionizable lipid nanoparticles
- PMID: 33786376
- PMCID: PMC7995196
- DOI: 10.1002/btm2.10213
Cytosolic delivery of nucleic acids: The case of ionizable lipid nanoparticles
Abstract
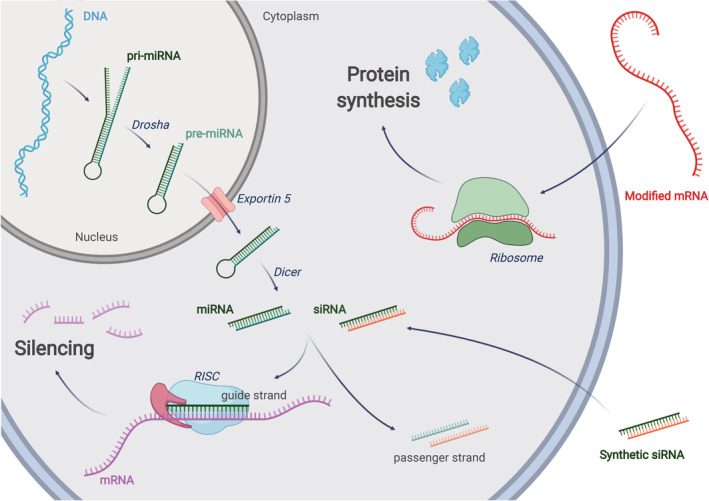
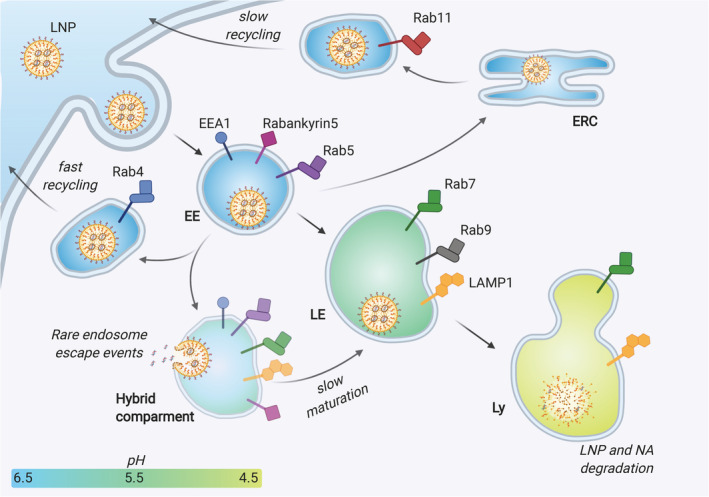
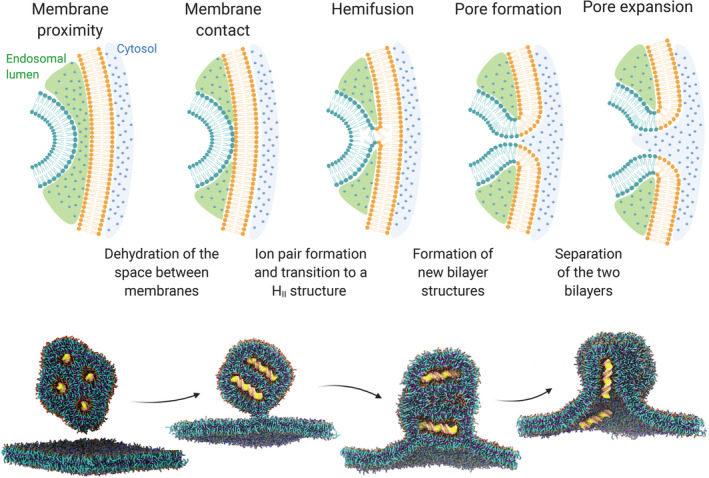
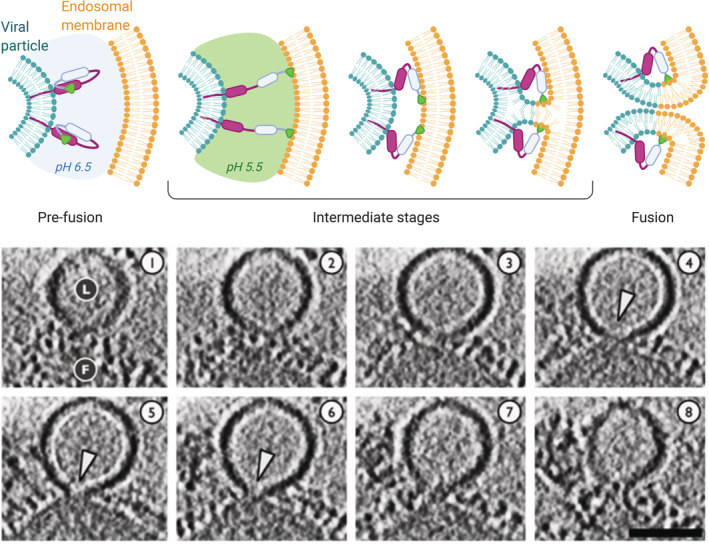
Ionizable lipid nanoparticles (LNPs) are the most clinically advanced nano-delivery system for therapeutic nucleic acids. The great effort put in the development of ionizable lipids with increased in vivo potency brought LNPs from the laboratory benches to the FDA approval of patisiran in 2018 and the ongoing clinical trials for mRNA-based vaccines against SARS-CoV-2. Despite these success stories, several challenges remain in RNA delivery, including what is known as "endosomal escape." Reaching the cytosol is mandatory for unleashing the therapeutic activity of RNA molecules, as their accumulation in other intracellular compartments would simply result in efficacy loss. In LNPs, the ability of ionizable lipids to form destabilizing non-bilayer structures at acidic pH is recognized as the key for endosomal escape and RNA cytosolic delivery. This is motivating a surge in studies aiming at designing novel ionizable lipids with improved biodegradation and safety profiles. In this work, we describe the journey of RNA-loaded LNPs across multiple intracellular barriers, from the extracellular space to the cytosol. In silico molecular dynamics modeling, in vitro high-resolution microscopy analyses, and in vivo imaging data are systematically reviewed to distill out the regulating mechanisms underlying the endosomal escape of RNA. Finally, a comparison with strategies employed by enveloped viruses to deliver their genetic material into cells is also presented. The combination of a multidisciplinary analytical toolkit for endosomal escape quantification and a nature-inspired design could foster the development of future LNPs with improved cytosolic delivery of nucleic acids.
Keywords: LNPs; RNA delivery; endosomal escape; intracellular delivery; ionizable lipids; mRNA; siRNA.
© 2021 The Authors. Bioengineering & Translational Medicine published by Wiley Periodicals LLC on behalf of The American Institute of Chemical Engineers.
Figures

References
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans Heterochronic gene lin‐4 encodes small RNAs with antisense complementarity to lin‐14. Cell. 1993;75:843‐854. - PubMed
-
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double‐stranded RNA in Caenorhabditis elegans . Nature. 1998;391:806‐811. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
