

Staphylococcus aureus cell wall structure and dynamics during host-pathogen interaction
- PMID: 33788901
- PMCID: PMC8041196
- DOI: 10.1371/journal.ppat.1009468
Staphylococcus aureus cell wall structure and dynamics during host-pathogen interaction
Abstract
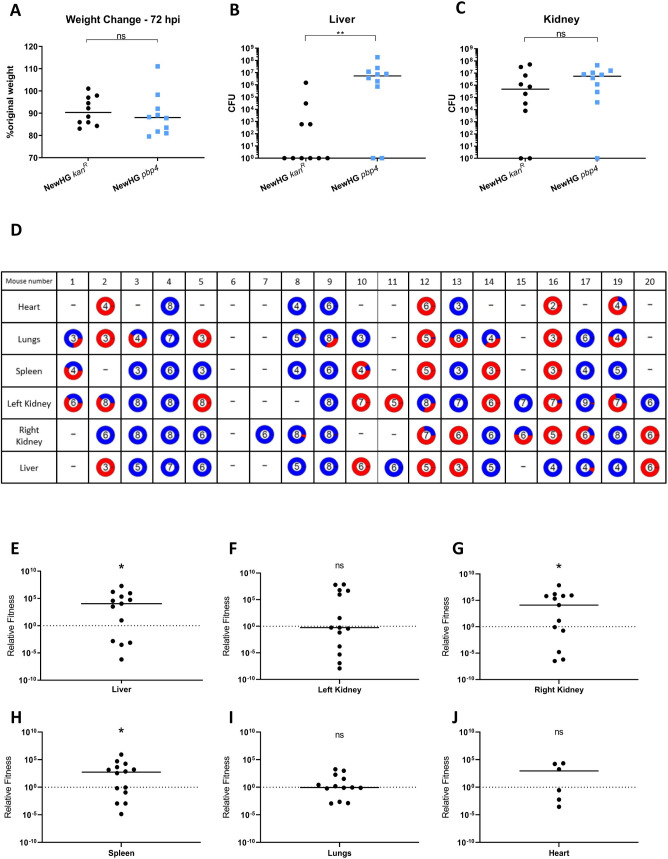
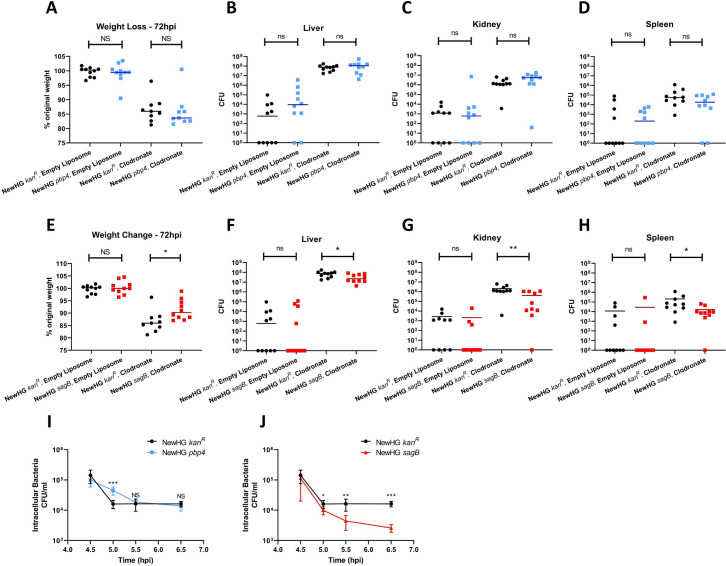
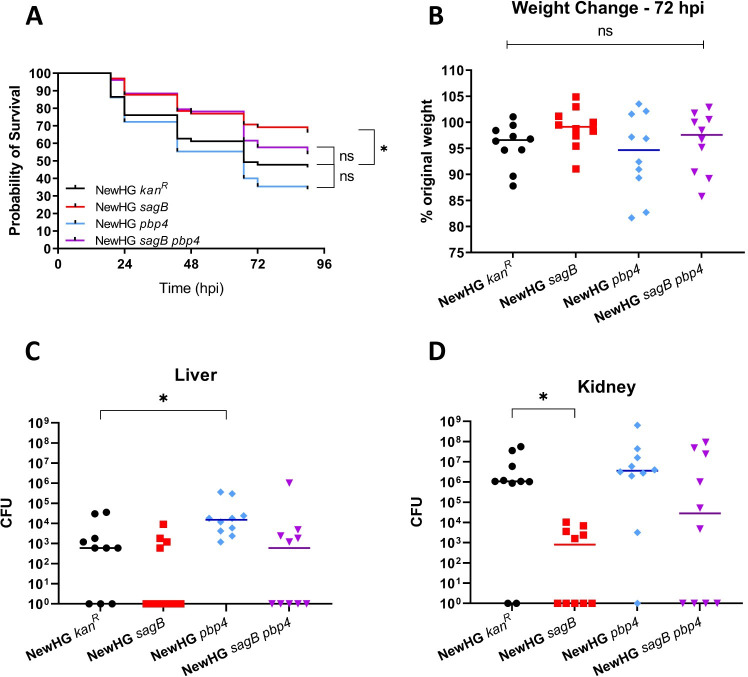
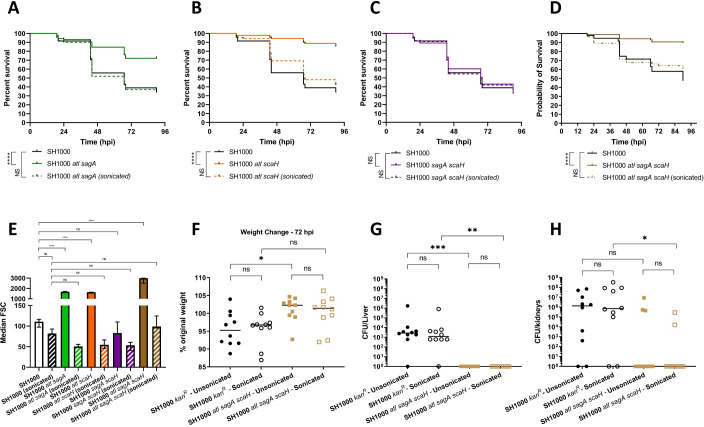
Peptidoglycan is the major structural component of the Staphylococcus aureus cell wall, in which it maintains cellular integrity, is the interface with the host, and its synthesis is targeted by some of the most crucial antibiotics developed. Despite this importance, and the wealth of data from in vitro studies, we do not understand the structure and dynamics of peptidoglycan during infection. In this study we have developed methods to harvest bacteria from an active infection in order to purify cell walls for biochemical analysis ex vivo. Isolated ex vivo bacterial cells are smaller than those actively growing in vitro, with thickened cell walls and reduced peptidoglycan crosslinking, similar to that of stationary phase cells. These features suggested a role for specific peptidoglycan homeostatic mechanisms in disease. As S. aureus missing penicillin binding protein 4 (PBP4) has reduced peptidoglycan crosslinking in vitro its role during infection was established. Loss of PBP4 resulted in an increased recovery of S. aureus from the livers of infected mice, which coincided with enhanced fitness within murine and human macrophages. Thicker cell walls correlate with reduced activity of peptidoglycan hydrolases. S. aureus has a family of 4 putative glucosaminidases, that are collectively crucial for growth. Loss of the major enzyme SagB, led to attenuation during murine infection and reduced survival in human macrophages. However, loss of the other three enzymes Atl, SagA and ScaH resulted in clustering dependent attenuation, in a zebrafish embryo, but not a murine, model of infection. A combination of pbp4 and sagB deficiencies resulted in a restoration of parental virulence. Our results, demonstrate the importance of appropriate cell wall structure and dynamics during pathogenesis, providing new insight to the mechanisms of disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Demonstration of the role of cell wall homeostasis in Staphylococcus aureus growth and the action of bactericidal antibiotics.Proc Natl Acad Sci U S A. 2021 Nov 2;118(44):e2106022118. doi: 10.1073/pnas.2106022118. Proc Natl Acad Sci U S A. 2021. PMID: 34716264 Free PMC article.
-
Bacterial Cell Enlargement Requires Control of Cell Wall Stiffness Mediated by Peptidoglycan Hydrolases.mBio. 2015 Jul 28;6(4):e00660. doi: 10.1128/mBio.00660-15. mBio. 2015. PMID: 26220963 Free PMC article.
-
PBP4 is required for serum-induced cell wall thickening and antibiotic tolerance in Staphylococcus aureus.Antimicrob Agents Chemother. 2024 Nov 6;68(11):e0096124. doi: 10.1128/aac.00961-24. Epub 2024 Oct 21. Antimicrob Agents Chemother. 2024. PMID: 39431816 Free PMC article.
-
Staphylococcus aureus cell wall maintenance - the multifaceted roles of peptidoglycan hydrolases in bacterial growth, fitness, and virulence.FEMS Microbiol Rev. 2022 Oct 28;46(5):fuac025. doi: 10.1093/femsre/fuac025. FEMS Microbiol Rev. 2022. PMID: 35675307 Free PMC article. Review.
-
The Staphylococcal Cell Wall.Microbiol Spectr. 2019 Jul;7(4):10.1128/microbiolspec.gpp3-0068-2019. doi: 10.1128/microbiolspec.GPP3-0068-2019. Microbiol Spectr. 2019. PMID: 31322105 Free PMC article. Review.
Cited by
-
Identification of a family of peptidoglycan transpeptidases reveals that Clostridioides difficile requires noncanonical cross-links for viability.Proc Natl Acad Sci U S A. 2024 Aug 20;121(34):e2408540121. doi: 10.1073/pnas.2408540121. Epub 2024 Aug 16. Proc Natl Acad Sci U S A. 2024. PMID: 39150786 Free PMC article.
-
Neutrophil extracellular traps in homeostasis and disease.Signal Transduct Target Ther. 2024 Sep 20;9(1):235. doi: 10.1038/s41392-024-01933-x. Signal Transduct Target Ther. 2024. PMID: 39300084 Free PMC article. Review.
-
Modulation of MRSA virulence gene expression by the wall teichoic acid enzyme TarO.Nat Commun. 2023 Mar 22;14(1):1594. doi: 10.1038/s41467-023-37310-5. Nat Commun. 2023. PMID: 36949052 Free PMC article.
-
The Cell Wall, Cell Membrane and Virulence Factors of Staphylococcus aureus and Their Role in Antibiotic Resistance.Microorganisms. 2023 Jan 19;11(2):259. doi: 10.3390/microorganisms11020259. Microorganisms. 2023. PMID: 36838224 Free PMC article. Review.
-
Phage lysins for intestinal microbiome modulation: current challenges and enabling techniques.Gut Microbes. 2024 Jan-Dec;16(1):2387144. doi: 10.1080/19490976.2024.2387144. Epub 2024 Aug 6. Gut Microbes. 2024. PMID: 39106212 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
