

Functional enhancer elements drive subclass-selective expression from mouse to primate neocortex
- PMID: 33789096
- PMCID: PMC8163032
- DOI: 10.1016/j.celrep.2021.108754
Functional enhancer elements drive subclass-selective expression from mouse to primate neocortex
Abstract
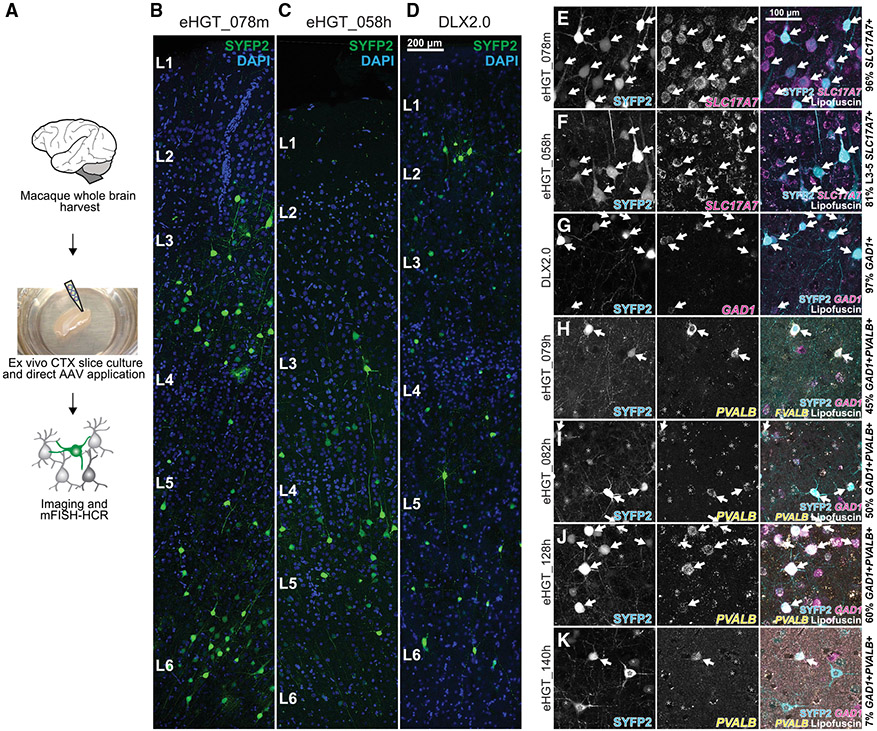
Viral genetic tools that target specific brain cell types could transform basic neuroscience and targeted gene therapy. Here, we use comparative open chromatin analysis to identify thousands of human-neocortical-subclass-specific putative enhancers from across the genome to control gene expression in adeno-associated virus (AAV) vectors. The cellular specificity of reporter expression from enhancer-AAVs is established by molecular profiling after systemic AAV delivery in mouse. Over 30% of enhancer-AAVs produce specific expression in the targeted subclass, including both excitatory and inhibitory subclasses. We present a collection of Parvalbumin (PVALB) enhancer-AAVs that show highly enriched expression not only in cortical PVALB cells but also in some subcortical PVALB populations. Five vectors maintain PVALB-enriched expression in primate neocortex. These results demonstrate how genome-wide open chromatin data mining and cross-species AAV validation can be used to create the next generation of non-species-restricted viral genetic tools.
Keywords: AAVs; ATAC-seq; brain cell types; enhancers; epigenetics; ex vivo brain slice; genetic tools; human; macaque; parvalbumin.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.K.M., L.T.G., E.E.H., H.Z., B.T., E.L., J.T.T., and B.P.L. are inventors on several U.S. patent applications related to this work. The remaining authors declare no competing interests.
Figures






References
-
- Anney RJL, Ripke S, Anttila V, Grove J, Holmans P, Huang H, Klei L, Lee PH, Medland SE, Neale B, et al.; Autism Spectrum Disorders Working Group of The Psychiatric Genomics Consortium (2017). Meta-analysis of GWAS of over 16,000 individuals with autism spectrum disorder highlights a novel locus at 10q24.32 and a significant overlap with schizophrenia. Mol. Autism 8, 21. - PMC - PubMed
-
- Autism Spectrum Disorder Working Group of the Psychiatry Genomics Consortium (2015). Dataset: PGC-ASD summary statistics from a meta-analysis of 5,305 ASD-diagnosed cases and 5,305 pseudocontrols of European descent (based on similarity to CEPH reference genotypes). http://www.med.unc.edu/pgc/results-and-downloads.
-
- Bakken TE, Jorstad NL, Hu Q, Lake BB, Tian W, Kalmbach BE, Crow M, Hodge RD, Krienen FM, Sorensen SA, et al. (2020). Evolution of cellular diversity in primary motor cortex of human, marmoset monkey, and mouse. BioRxiv, 2020.03.31.016972.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
