

The Balance of MU-Opioid, Dopamine D2 and Adenosine A2A Heteroreceptor Complexes in the Ventral Striatal-Pallidal GABA Antireward Neurons May Have a Significant Role in Morphine and Cocaine Use Disorders
- PMID: 33790790
- PMCID: PMC8005530
- DOI: 10.3389/fphar.2021.627032
The Balance of MU-Opioid, Dopamine D2 and Adenosine A2A Heteroreceptor Complexes in the Ventral Striatal-Pallidal GABA Antireward Neurons May Have a Significant Role in Morphine and Cocaine Use Disorders
Abstract
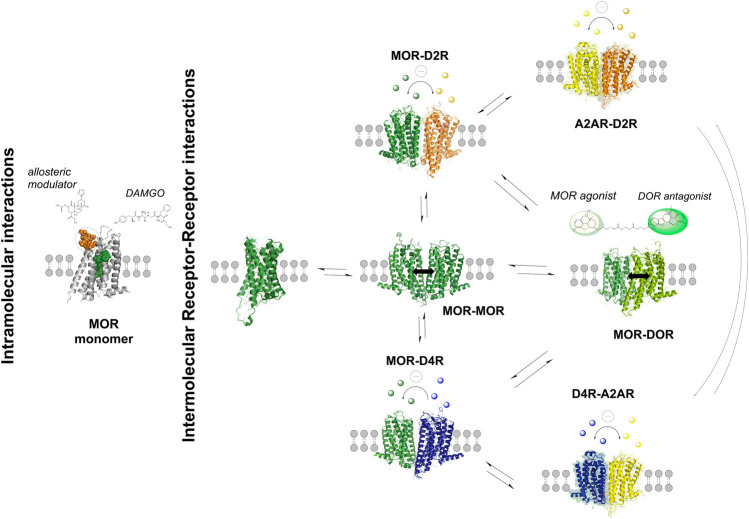
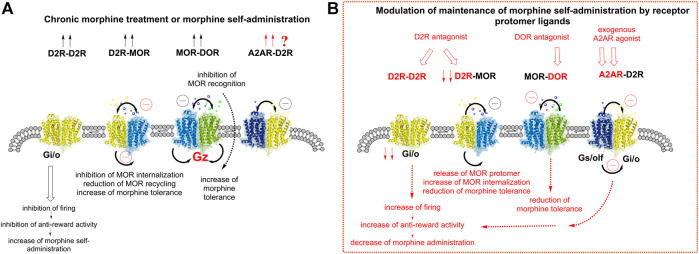
The widespread distribution of heteroreceptor complexes with allosteric receptor-receptor interactions in the CNS represents a novel integrative molecular mechanism in the plasma membrane of neurons and glial cells. It was proposed that they form the molecular basis for learning and short-and long-term memories. This is also true for drug memories formed during the development of substance use disorders like morphine and cocaine use disorders. In cocaine use disorder it was found that irreversible A2AR-D2R complexes with an allosteric brake on D2R recognition and signaling are formed in increased densities in the ventral enkephalin positive striatal-pallidal GABA antireward neurons. In this perspective article we discuss and propose how an increase in opioid heteroreceptor complexes, containing MOR-DOR, MOR-MOR and MOR-D2R, and their balance with each other and A2AR-D2R complexes in the striatal-pallidal enkephalin positive GABA antireward neurons, may represent markers for development of morphine use disorders. We suggest that increased formation of MOR-DOR complexes takes place in the striatal-pallidal enkephalin positive GABA antireward neurons after chronic morphine treatment in part through recruitment of MOR from the MOR-D2R complexes due to the possibility that MOR upon morphine treatment can develop a higher affinity for DOR. As a result, increased numbers of D2R monomers/homomers in these neurons become free to interact with the A2A receptors found in high densities within such neurons. Increased numbers of A2AR-D2R heteroreceptor complexes are formed and contribute to enhanced firing of these antireward neurons due to loss of inhibitory D2R protomer signaling which finally leads to the development of morphine use disorder. Development of cocaine use disorder may instead be reduced through enkephalin induced activation of the MOR-DOR complex inhibiting the activity of the enkephalin positive GABA antireward neurons. Altogether, we propose that these altered complexes could be pharmacological targets to modulate the reward and the development of substance use disorders.
Keywords: G protein-coupled receptor; adenosine A2A receptor; cocaine use disorder; dopamine D2 receptor; morphine use disorder; mu opioid receptor; oligomerization, morphine.
Copyright © 2021 Borroto-Escuela, Wydra, Fores-Pons, Vasudevan, Romero-Fernandez, Frankowska, Ferraro, Beggiato, Crespo-Ramirez, Rivera, Rocha, Perez de la Mora, Stove, Filip and Fuxe.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Borroto-Escuela D. O., Brito I., Di Palma M., Jiménez-Beristain A., Narvaez M., Corrales F., et al. (2015). On the role of the balance of GPCR Homo/heteroreceptor complexes in the brain. J. Adv. Neurosci. Res. 2, 36–44. 10.15379/2409-3564.2015.02.01.5 - DOI
-
- Borroto-Escuela D. O., Narvaez M., Wydra K., Pintsuk J., Pinton L., Jimenez-Beristain A., et al. (2017b). Cocaine self-administration specifically increases A2AR-D2R and D2R-sigma1R heteroreceptor complexes in the rat nucleus accumbens shell. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 155, 24–31. 10.1016/j.pbb.2017.03.003 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials