

Transport mechanisms at the malaria parasite-host cell interface
- PMID: 33793667
- PMCID: PMC8016102
- DOI: 10.1371/journal.ppat.1009394
Transport mechanisms at the malaria parasite-host cell interface
Abstract
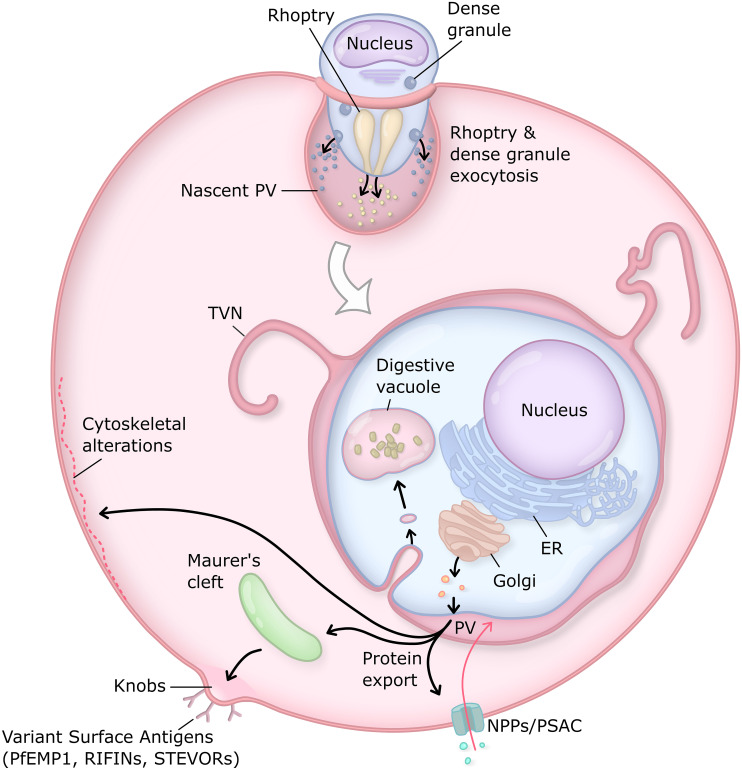
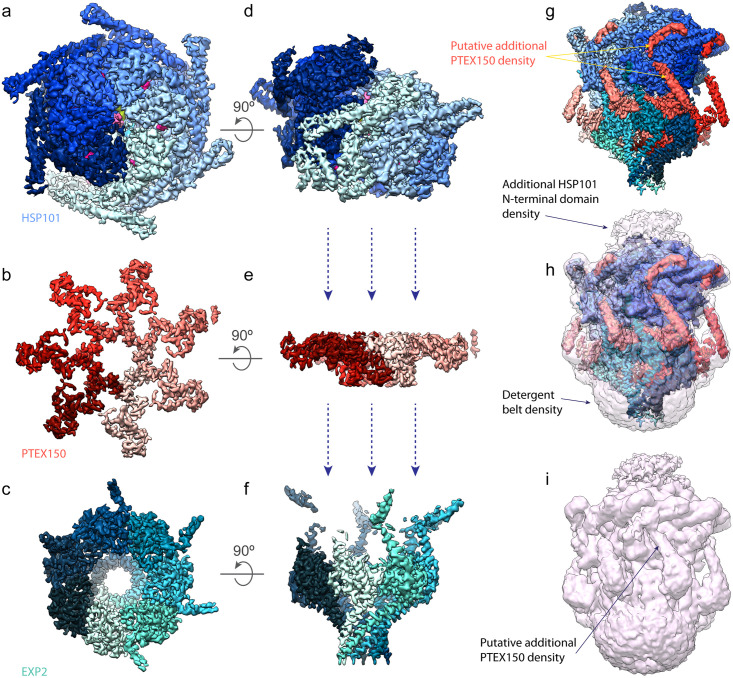
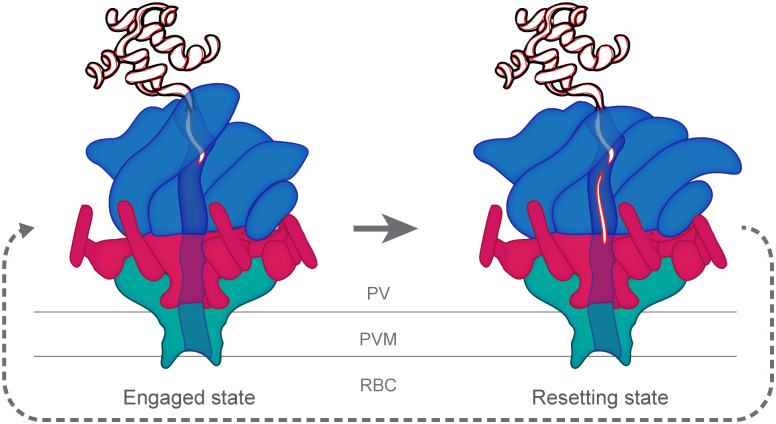
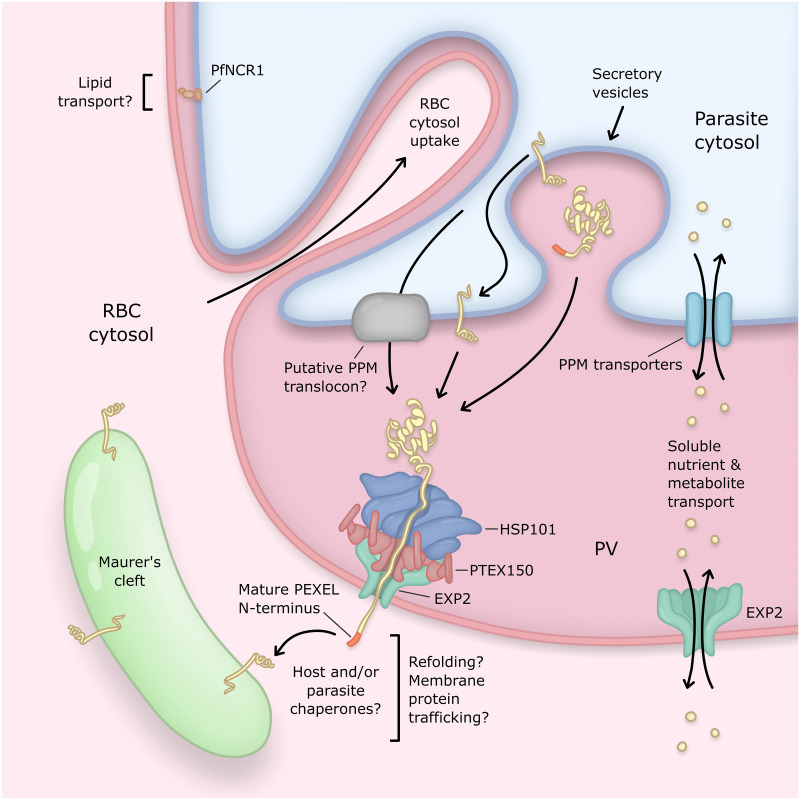
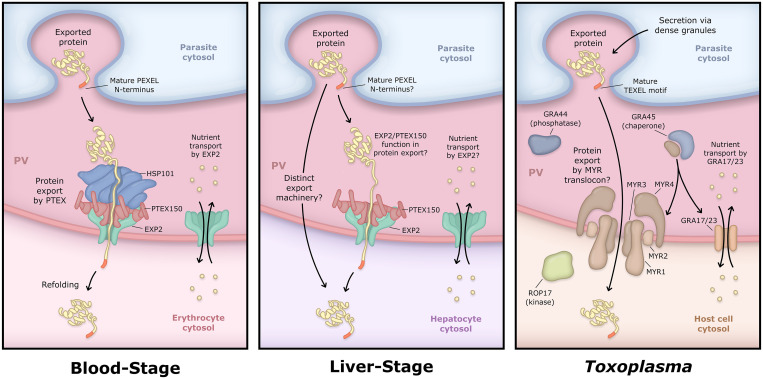
Obligate intracellular malaria parasites reside within a vacuolar compartment generated during invasion which is the principal interface between pathogen and host. To subvert their host cell and support their metabolism, these parasites coordinate a range of transport activities at this membrane interface that are critically important to parasite survival and virulence, including nutrient import, waste efflux, effector protein export, and uptake of host cell cytosol. Here, we review our current understanding of the transport mechanisms acting at the malaria parasite vacuole during the blood and liver-stages of development with a particular focus on recent advances in our understanding of effector protein translocation into the host cell by the Plasmodium Translocon of EXported proteins (PTEX) and small molecule transport by the PTEX membrane-spanning pore EXP2. Comparison to Toxoplasma gondii and other related apicomplexans is provided to highlight how similar and divergent mechanisms are employed to fulfill analogous transport activities.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Lingelbach K, Joiner KA. The parasitophorous vacuole membrane surrounding Plasmodium and Toxoplasma: an unusual compartment in infected cells. J Cell Sci. 1998;111(Pt 11):1467–75. . - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
