

Redox regulation of chloroplast metabolism
- PMID: 33793865
- PMCID: PMC8154093
- DOI: 10.1093/plphys/kiaa062
Redox regulation of chloroplast metabolism
Abstract
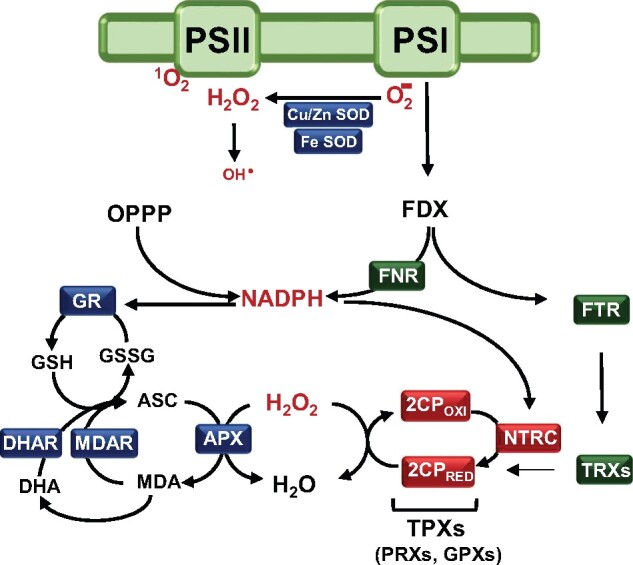
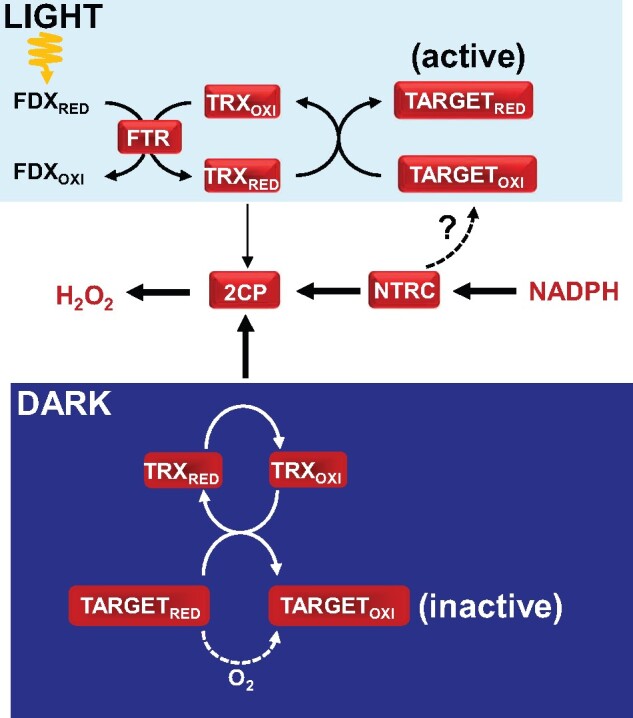
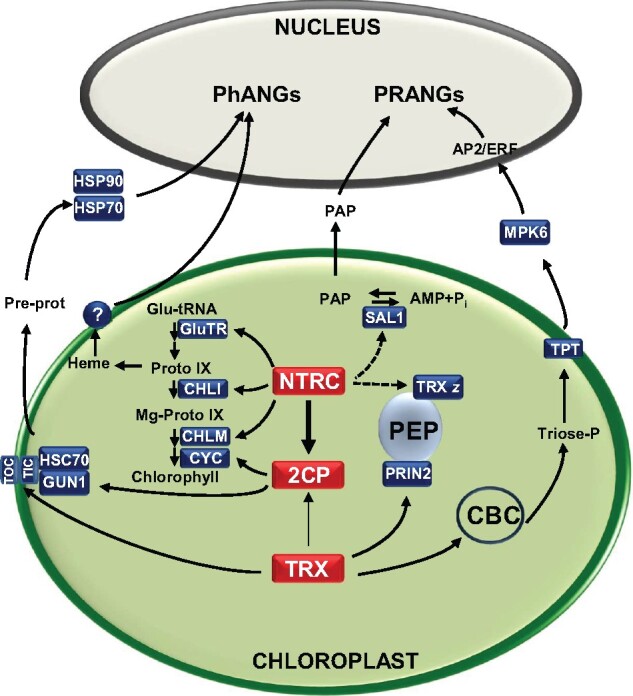
Regulation of enzyme activity based on thiol-disulfide exchange is a regulatory mechanism in which the protein disulfide reductase activity of thioredoxins (TRXs) plays a central role. Plant chloroplasts are equipped with a complex set of up to 20 TRXs and TRX-like proteins, the activity of which is supported by reducing power provided by photosynthetically reduced ferredoxin (FDX) with the participation of a FDX-dependent TRX reductase (FTR). Therefore, the FDX-FTR-TRXs pathway allows the regulation of redox-sensitive chloroplast enzymes in response to light. In addition, chloroplasts contain an NADPH-dependent redox system, termed NTRC, which allows the use of NADPH in the redox network of these organelles. Genetic approaches using mutants of Arabidopsis (Arabidopsis thaliana) in combination with biochemical and physiological studies have shown that both redox systems, NTRC and FDX-FTR-TRXs, participate in fine-tuning chloroplast performance in response to changes in light intensity. Moreover, these studies revealed the participation of 2-Cys peroxiredoxin (2-Cys PRX), a thiol-dependent peroxidase, in the control of the reducing activity of chloroplast TRXs as well as in the rapid oxidation of stromal enzymes upon darkness. In this review, we provide an update on recent findings regarding the redox regulatory network of plant chloroplasts, focusing on the functional relationship of 2-Cys PRXs with NTRC and the FDX-FTR-TRXs redox systems for fine-tuning chloroplast performance in response to changes in light intensity and darkness. Finally, we consider redox regulation as an additional layer of control of the signaling function of the chloroplast.
© The Author(s) 2020. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
References
-
- Alkhalfioui F, Renard M, Montrichard F (2007) Unique properties of NADP-thioredoxin reductase C in legumes. J Exp Bot 58:969–978 - PubMed
-
- Arsova B, Hoja U, Wimmelbacher M, Greiner E, Ustun S, Melzer M, Petersen K, Lein W, Börnke F (2010) Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 22:1498–1515 - PMC - PubMed
-
- Awad J, Stotz HU, Fekete A, Krischke M, Engert C, Havaux M, Berger S, Mueller MJ (2015) 2-Cysteine peroxiredoxins and thylakoid ascorbate peroxidase create a water–water cycle that is essential to protect the photosynthetic apparatus under high light stress conditions. Plant Physiol 167:1592–1603 - PMC - PubMed
