

Plasma membrane phylloquinone biosynthesis in nonphotosynthetic parasitic plants
- PMID: 33793953
- PMCID: PMC8133638
- DOI: 10.1093/plphys/kiab031
Plasma membrane phylloquinone biosynthesis in nonphotosynthetic parasitic plants
Abstract
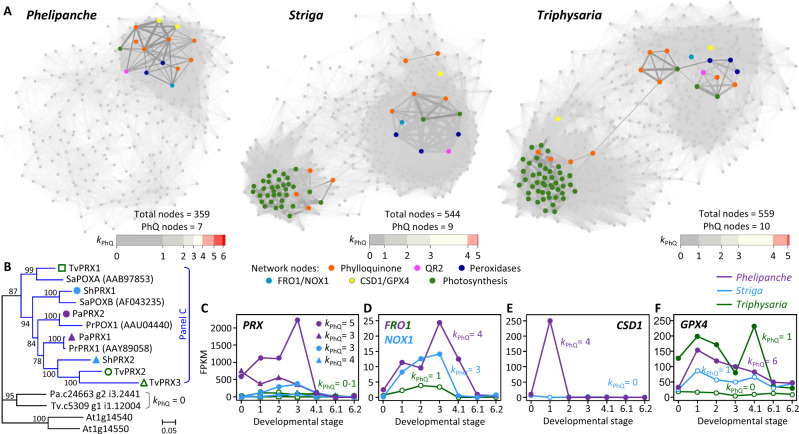
Nonphotosynthetic holoparasites exploit flexible targeting of phylloquinone biosynthesis to facilitate plasma membrane redox signaling. Phylloquinone is a lipophilic naphthoquinone found predominantly in chloroplasts and best known for its function in photosystem I electron transport and disulfide bridge formation of photosystem II subunits. Phylloquinone has also been detected in plasma membrane (PM) preparations of heterotrophic tissues with potential transmembrane redox function, but the molecular basis for this noncanonical pathway is unknown. Here, we provide evidence of PM phylloquinone biosynthesis in a nonphotosynthetic holoparasite Phelipanche aegyptiaca. A nonphotosynthetic and nonplastidial role for phylloquinone is supported by transcription of phylloquinone biosynthetic genes during seed germination and haustorium development, by PM-localization of alternative terminal enzymes, and by detection of phylloquinone in germinated seeds. Comparative gene network analysis with photosynthetically competent parasites revealed a bias of P. aegyptiaca phylloquinone genes toward coexpression with oxidoreductases involved in PM electron transport. Genes encoding the PM phylloquinone pathway are also present in several photoautotrophic taxa of Asterids, suggesting an ancient origin of multifunctionality. Our findings suggest that nonphotosynthetic holoparasites exploit alternative targeting of phylloquinone for transmembrane redox signaling associated with parasitism.
© The Author(s) 2021. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures




References
-
- Alexa A, Rahnenführer J, Lengauer T (2006) Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 22:1600–1607 - PubMed
-
- Attacha S, Solbach D, Bela K, Moseler A, Wagner S, Schwarzländer M, Aller I, Müller Stefanie J, Meyer Andreas J (2017) Glutathione peroxidase‐like enzymes cover five distinct cell compartments and membrane surfaces in Arabidopsis thaliana. Plant Cell Environ 40:1281–1295 - PubMed
-
- Babujee L, Wurtz V, Ma C, Lueder F, Soni P, van Dorsselaer A, Reumann S (2010) The proteome map of spinach leaf peroxisomes indicates partial compartmentalization of phylloquinone (vitamin K1) biosynthesis in plant peroxisomes. J Exp Bot 61:1441–1453 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
