

Gamma rhythm communication between entorhinal cortex and dentate gyrus neuronal assemblies
- PMID: 33795429
- PMCID: PMC8285088
- DOI: 10.1126/science.abf3119
Gamma rhythm communication between entorhinal cortex and dentate gyrus neuronal assemblies
Abstract
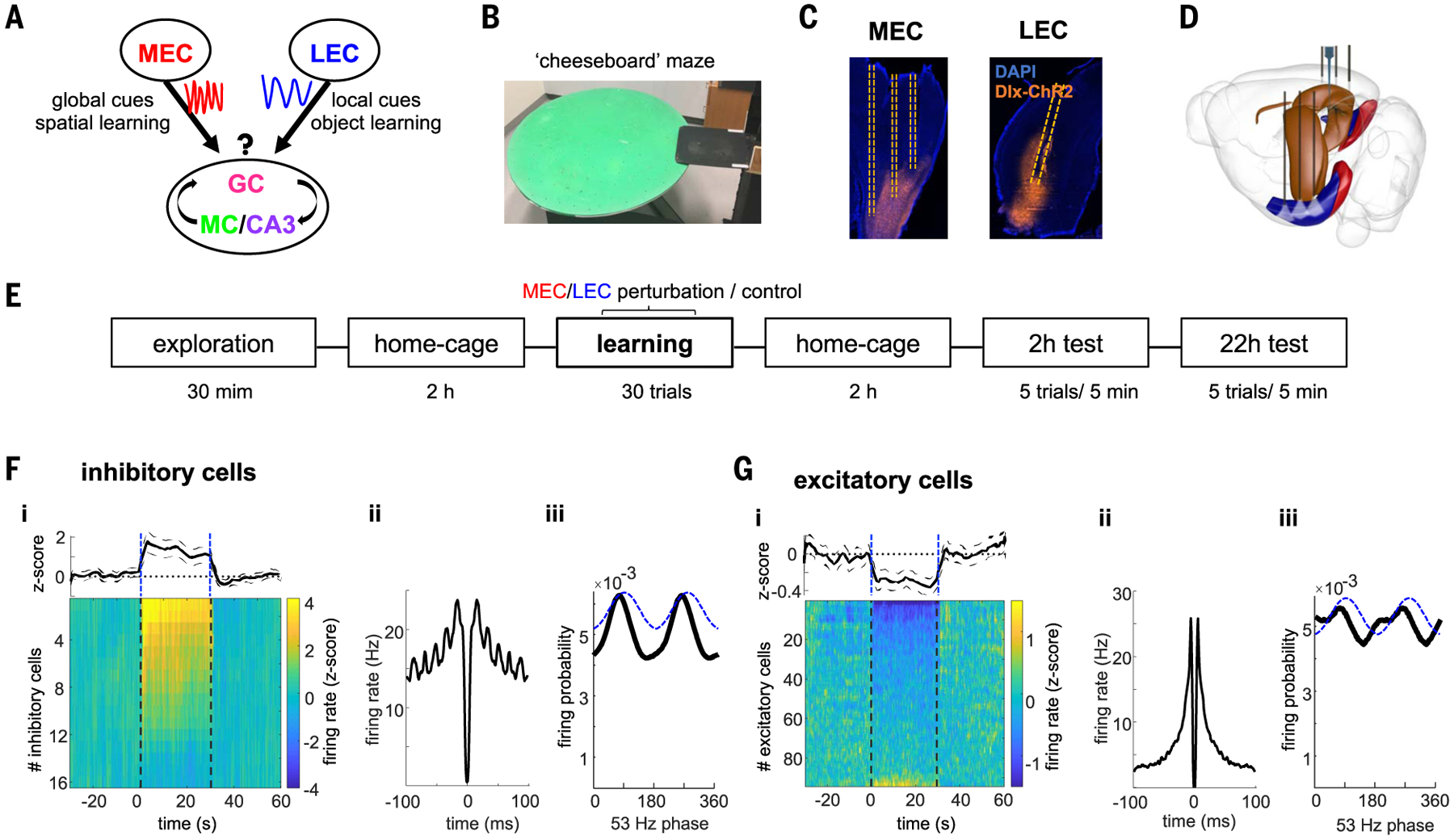
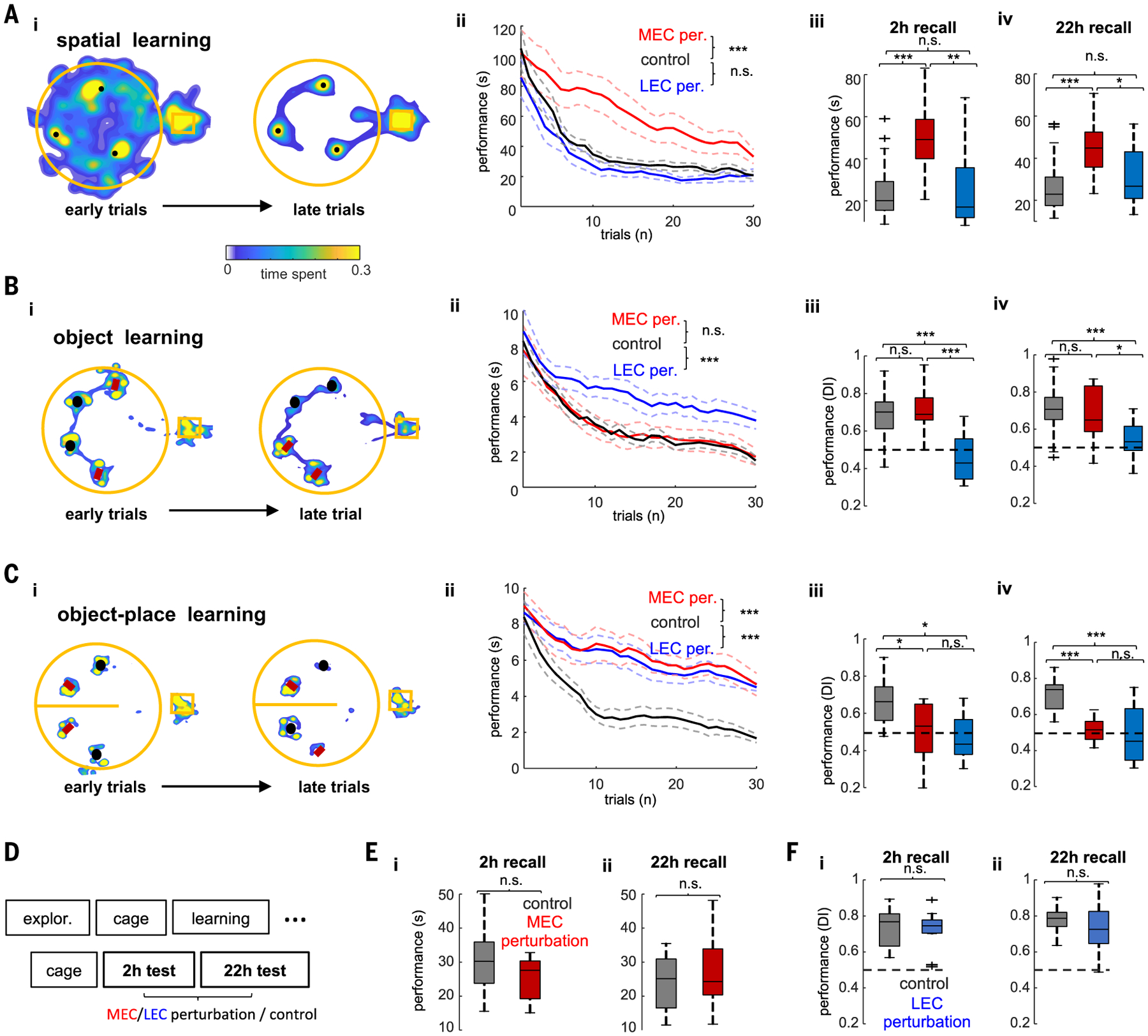
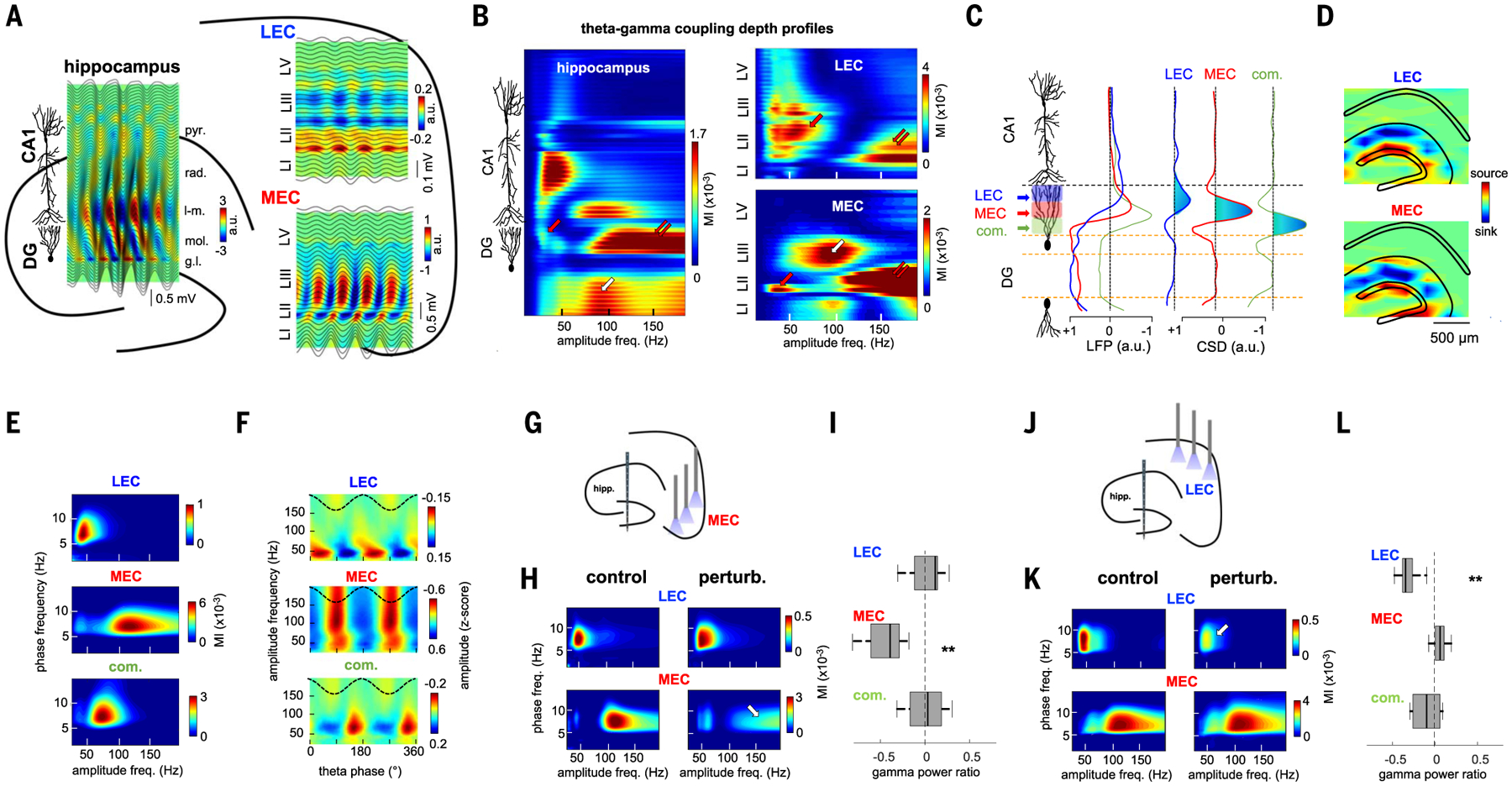
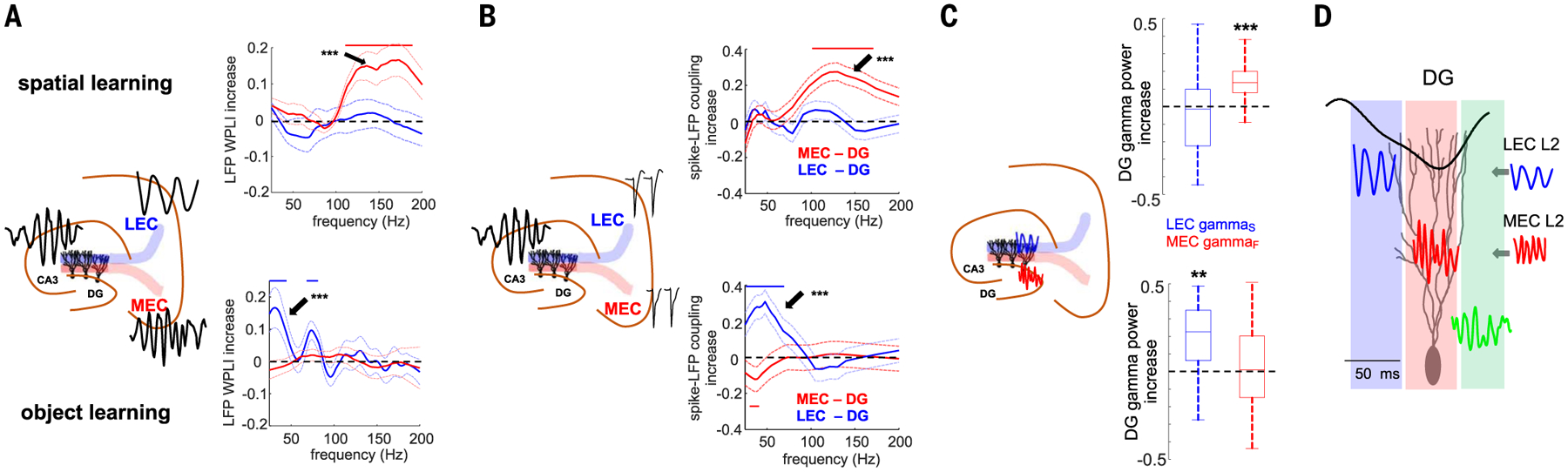
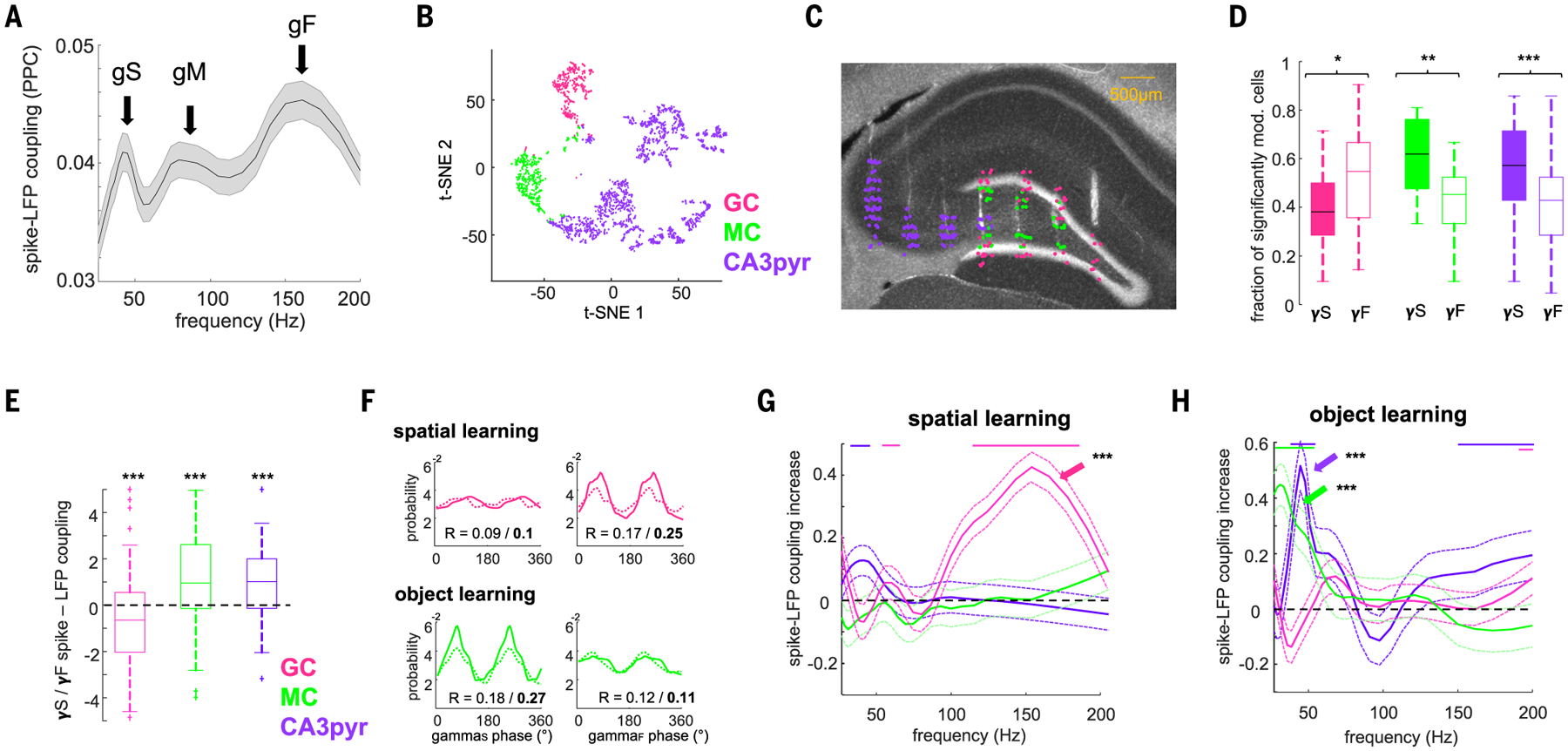
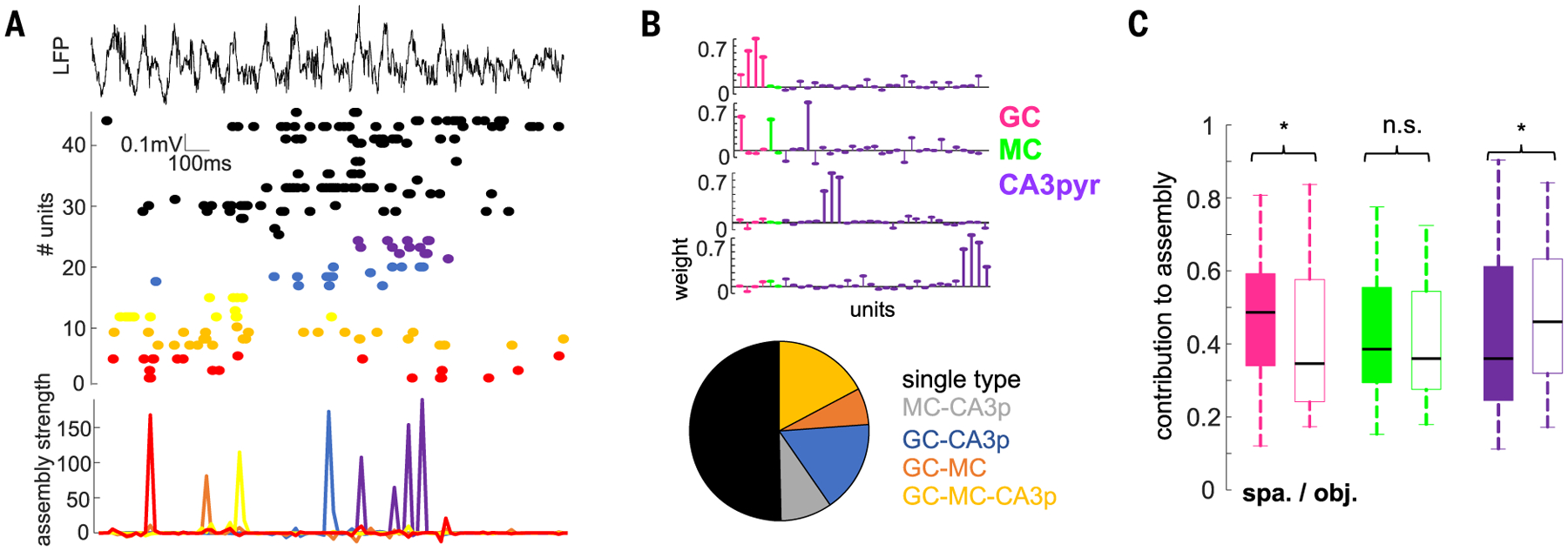
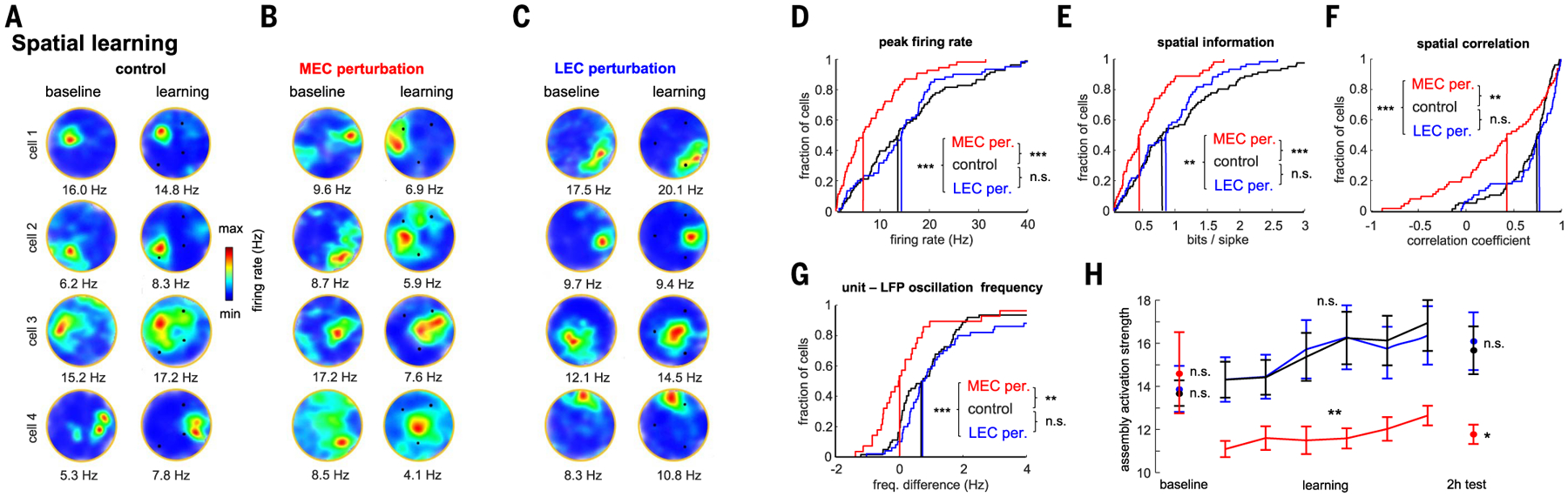
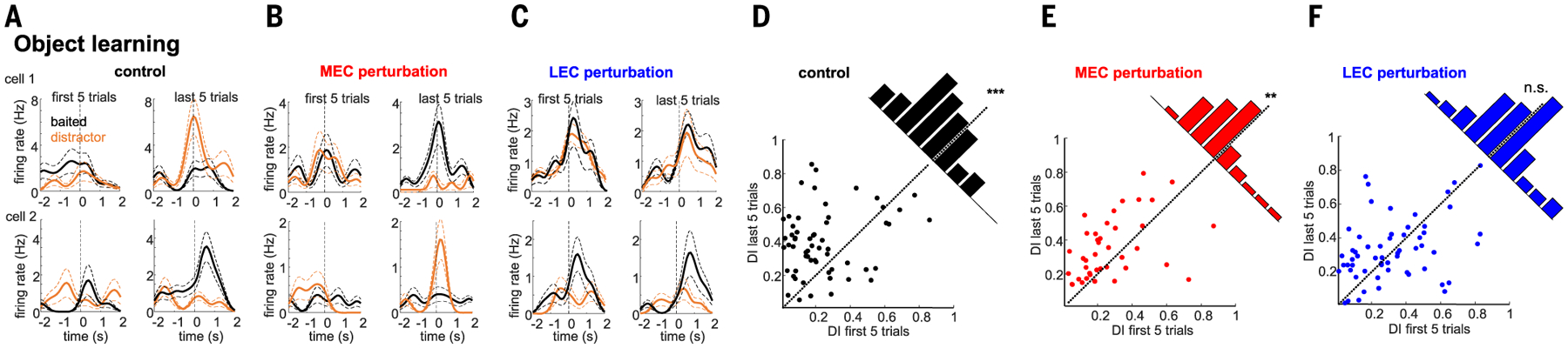
Gamma oscillations are thought to coordinate the spike timing of functionally specialized neuronal ensembles across brain regions. To test this hypothesis, we optogenetically perturbed gamma spike timing in the rat medial (MEC) and lateral (LEC) entorhinal cortices and found impairments in spatial and object learning tasks, respectively. MEC and LEC were synchronized with the hippocampal dentate gyrus through high- and low-gamma-frequency rhythms, respectively, and engaged either granule cells or mossy cells and CA3 pyramidal cells in a task-dependent manner. Gamma perturbation disrupted the learning-induced assembly organization of target neurons. Our findings imply that pathway-specific gamma oscillations route task-relevant information between distinct neuronal subpopulations in the entorhinal-hippocampal circuit. We hypothesize that interregional gamma-time-scale spike coordination is a mechanism of neuronal communication.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
