

Association of structural variation with cardiometabolic traits in Finns
- PMID: 33798444
- PMCID: PMC8059371
- DOI: 10.1016/j.ajhg.2021.03.008
Association of structural variation with cardiometabolic traits in Finns
Abstract
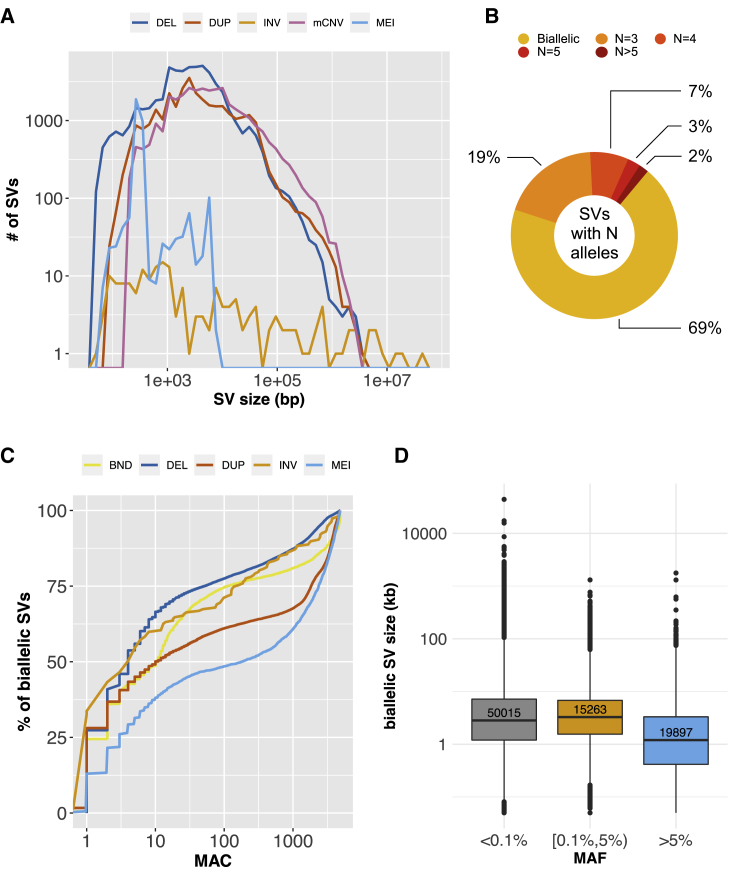
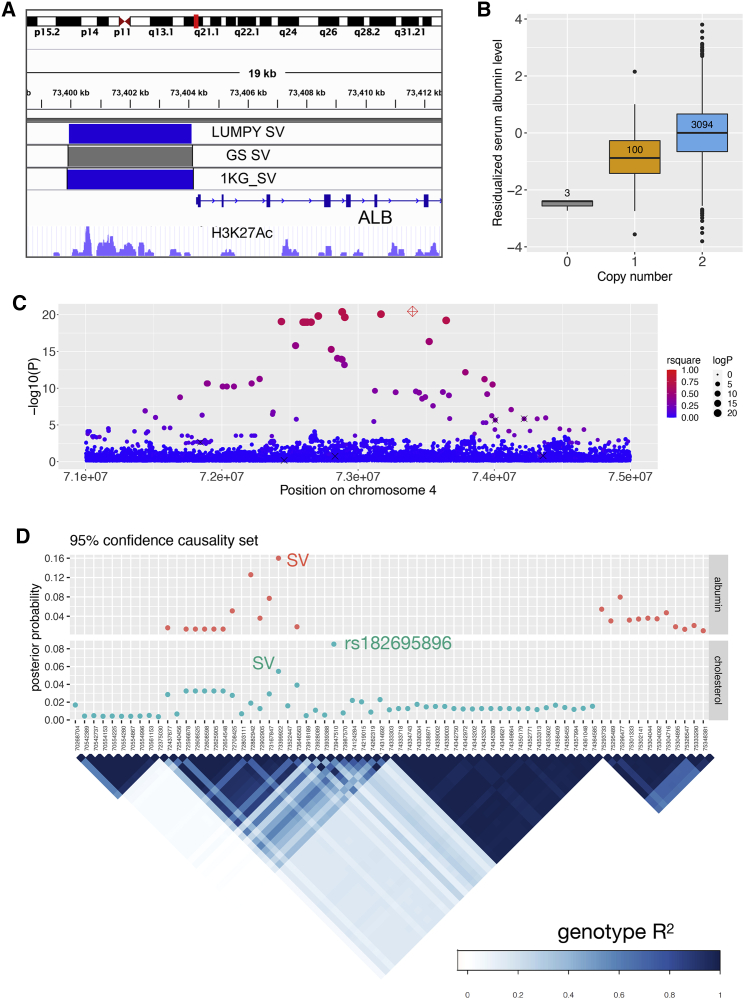
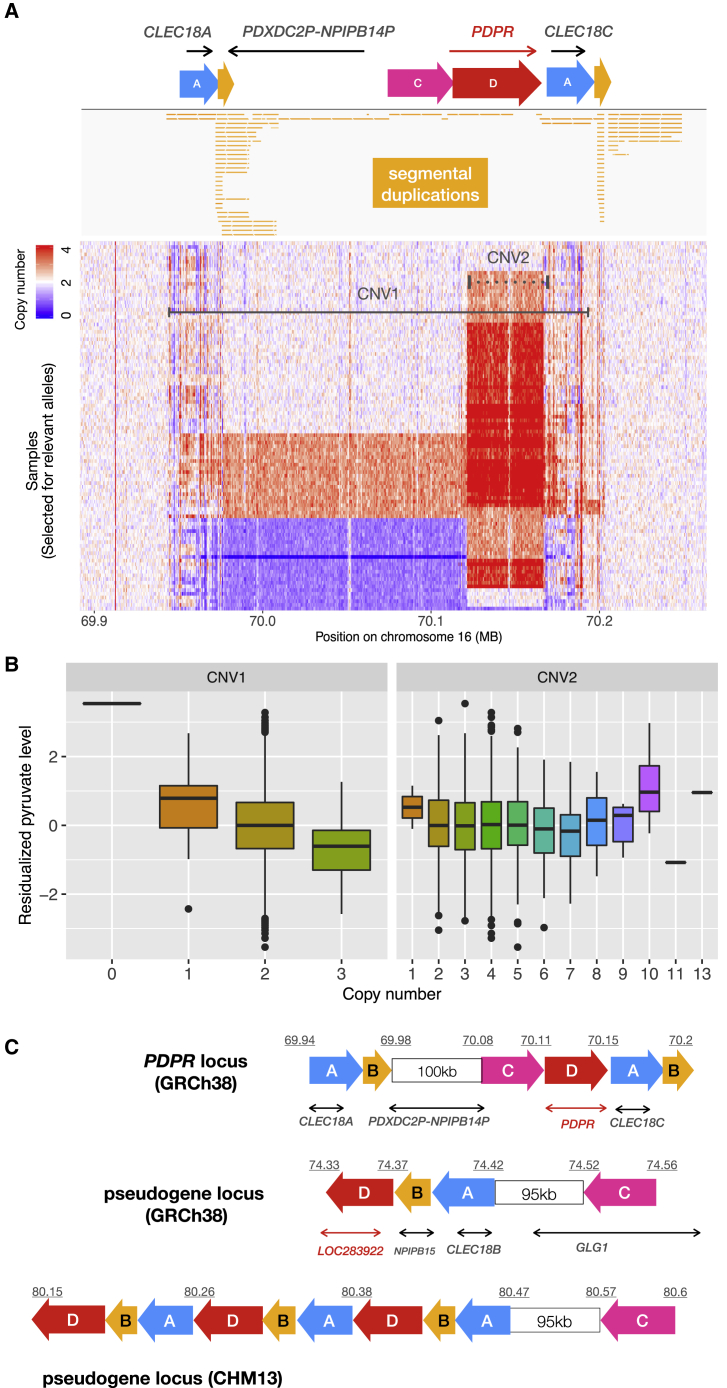
The contribution of genome structural variation (SV) to quantitative traits associated with cardiometabolic diseases remains largely unknown. Here, we present the results of a study examining genetic association between SVs and cardiometabolic traits in the Finnish population. We used sensitive methods to identify and genotype 129,166 high-confidence SVs from deep whole-genome sequencing (WGS) data of 4,848 individuals. We tested the 64,572 common and low-frequency SVs for association with 116 quantitative traits and tested candidate associations using exome sequencing and array genotype data from an additional 15,205 individuals. We discovered 31 genome-wide significant associations at 15 loci, including 2 loci at which SVs have strong phenotypic effects: (1) a deletion of the ALB promoter that is greatly enriched in the Finnish population and causes decreased serum albumin level in carriers (p = 1.47 × 10-54) and is also associated with increased levels of total cholesterol (p = 1.22 × 10-28) and 14 additional cholesterol-related traits, and (2) a multi-allelic copy number variant (CNV) at PDPR that is strongly associated with pyruvate (p = 4.81 × 10-21) and alanine (p = 6.14 × 10-12) levels and resides within a structurally complex genomic region that has accumulated many rearrangements over evolutionary time. We also confirmed six previously reported associations, including five led by stronger signals in single nucleotide variants (SNVs) and one linking recurrent HP gene deletion and cholesterol levels (p = 6.24 × 10-10), which was also found to be strongly associated with increased glycoprotein level (p = 3.53 × 10-35). Our study confirms that integrating SVs in trait-mapping studies will expand our knowledge of genetic factors underlying disease risk.
Keywords: Finnish population; cardiometabolic traits; genome-wide association study; structural variation.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
N.O.S. has received research funding from Regeneron Pharmaceuticals unrelated to this study. The rest of the authors declare no competing interests.
Figures
References
-
- Ortega F.B., Lavie C.J., Blair S.N. Obesity and Cardiovascular Disease. Circ. Res. 2016;118:1752–1770. - PubMed
-
- Kolifarhood G., Daneshpour M., Hadaegh F., Sabour S., Mozafar Saadati H., Akbar Haghdoust A., Akbarzadeh M., Sedaghati-Khayat B., Khosravi N. Heritability of blood pressure traits in diverse populations: a systematic review and meta-analysis. J. Hum. Hypertens. 2019;33:775–785. - PubMed
-
- Campbell Am L.V. Genetics of obesity. Aust. Fam. Physician. 2017;46:456–459. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
