

Molecular Mechanisms for Bacterial Potassium Homeostasis
- PMID: 33798529
- PMCID: PMC9041122
- DOI: 10.1016/j.jmb.2021.166968
Molecular Mechanisms for Bacterial Potassium Homeostasis
Abstract
Potassium ion homeostasis is essential for bacterial survival, playing roles in osmoregulation, pH homeostasis, regulation of protein synthesis, enzyme activation, membrane potential adjustment and electrical signaling. To accomplish such diverse physiological tasks, it is not surprising that a single bacterium typically encodes several potassium uptake and release systems. To understand the role each individual protein fulfills and how these proteins work in concert, it is important to identify the molecular details of their function. One needs to understand whether the systems transport ions actively or passively, and what mechanisms or ligands lead to the activation or inactivation of individual systems. Combining mechanistic information with knowledge about the physiology under different stress situations, such as osmostress, pH stress or nutrient limitation, one can identify the task of each system and deduce how they are coordinated with each other. By reviewing the general principles of bacterial membrane physiology and describing the molecular architecture and function of several bacterial K+-transporting systems, we aim to provide a framework for microbiologists studying bacterial potassium homeostasis and the many K+-translocating systems that are still poorly understood.
Keywords: K(+) transport; bacterial physiology; membrane potential; principles of K(+) transporters and channels; structural biology.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
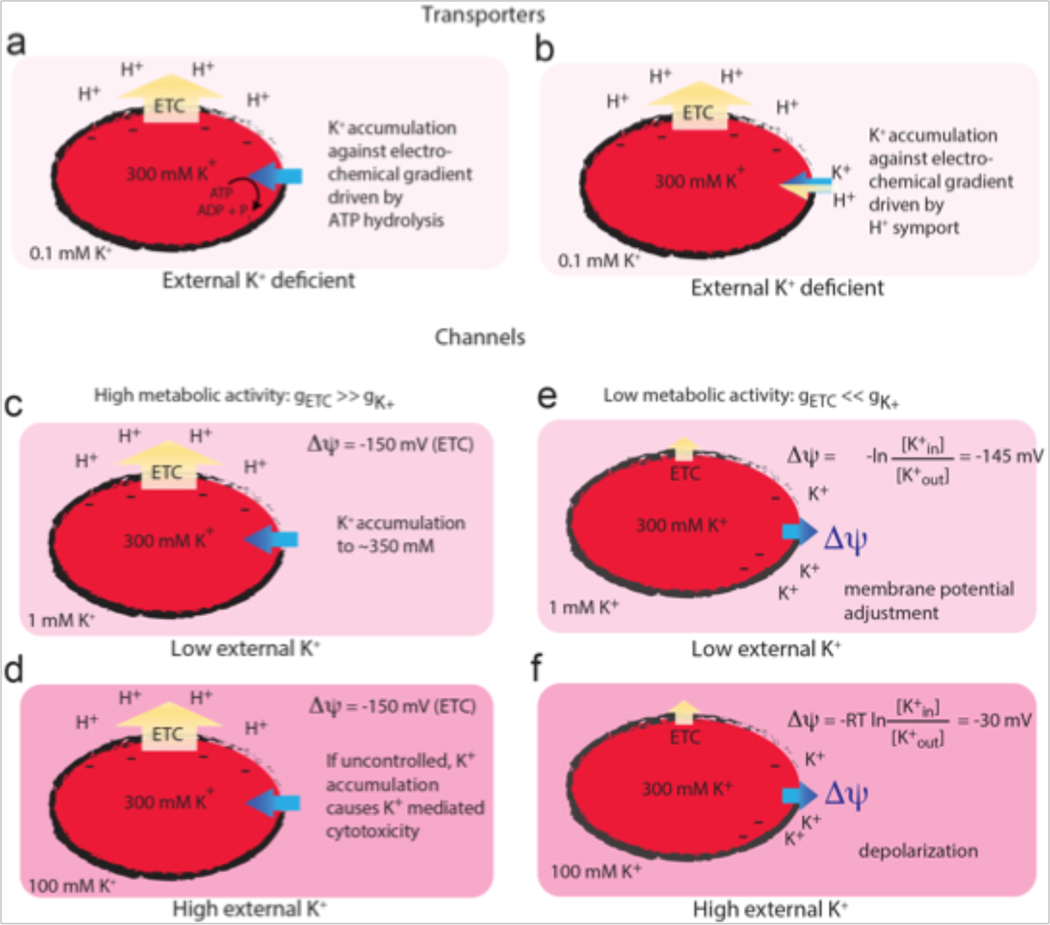
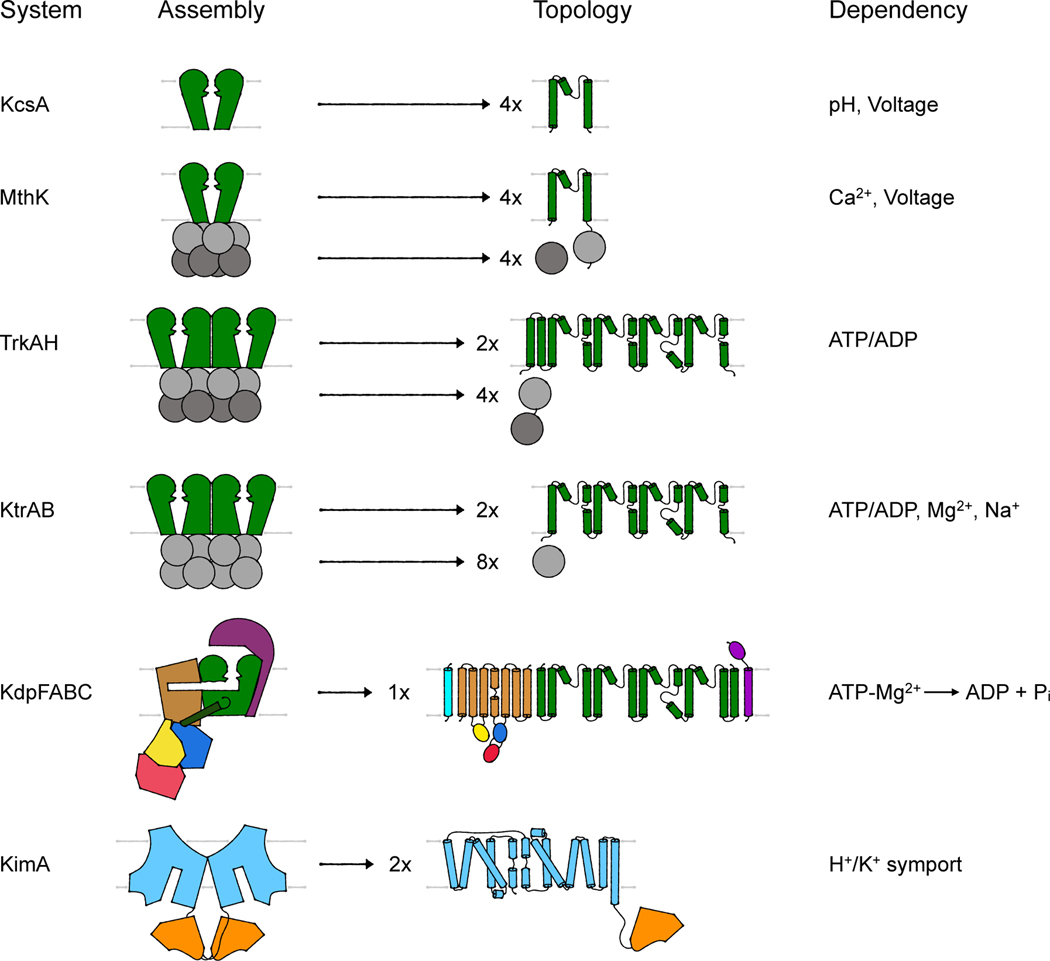
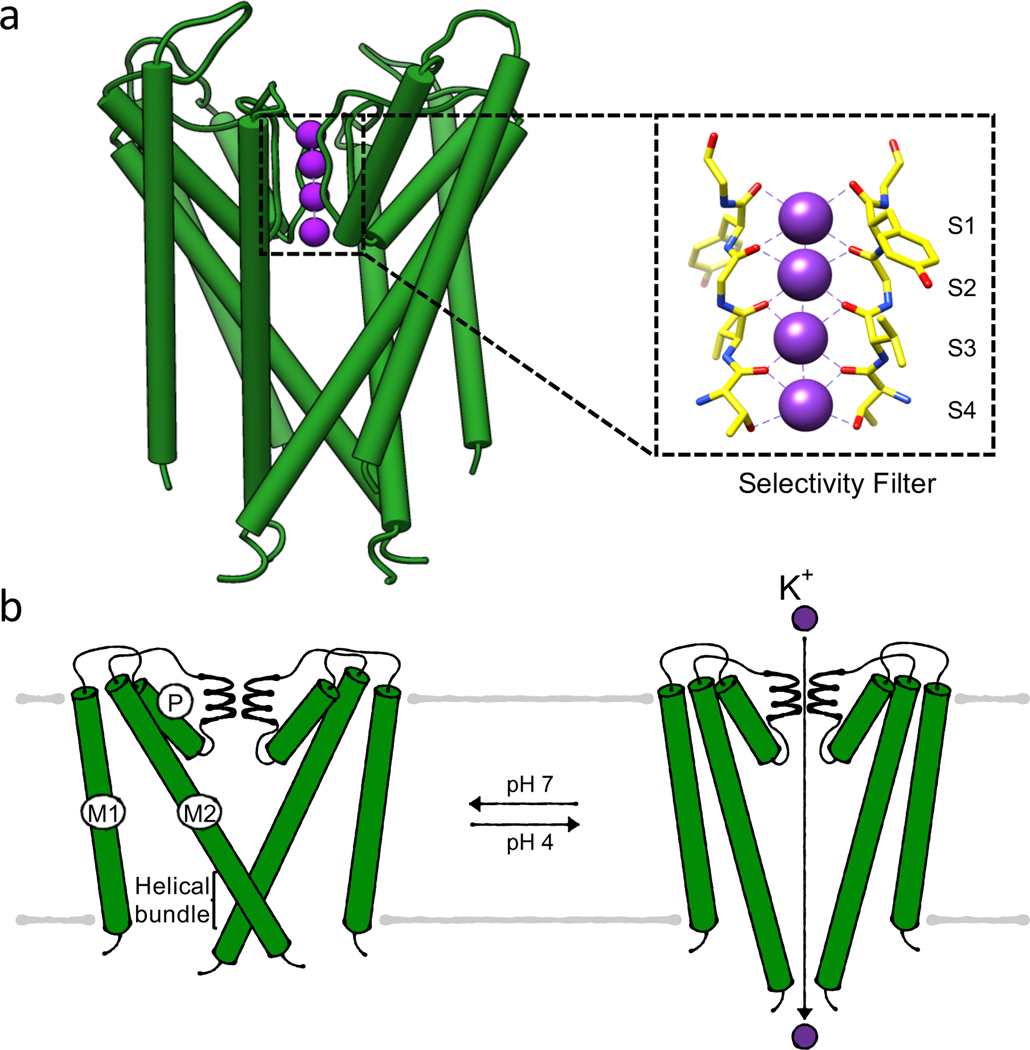
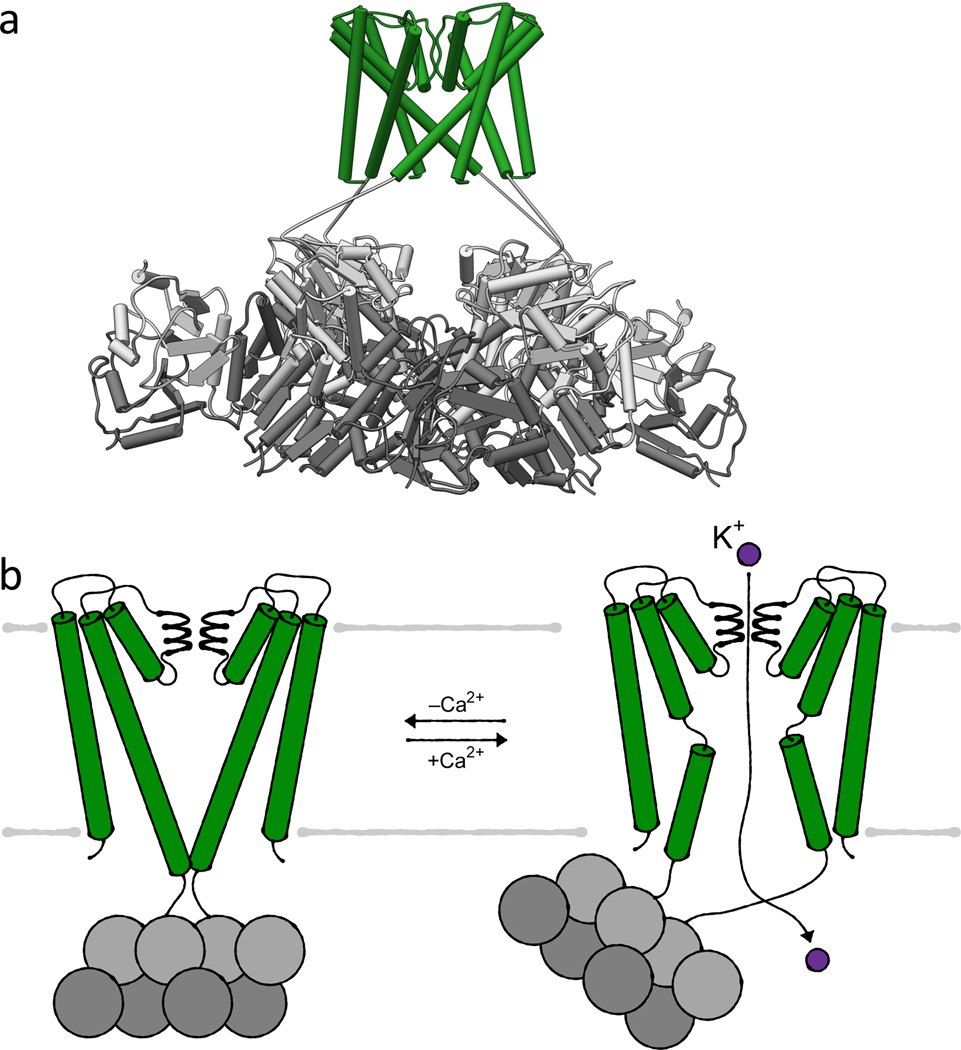
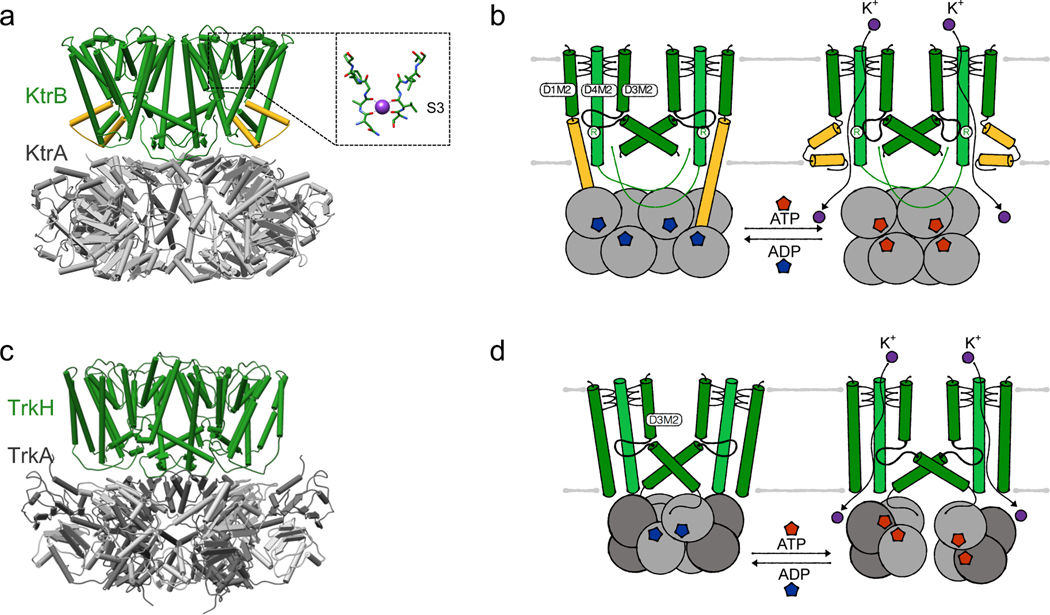
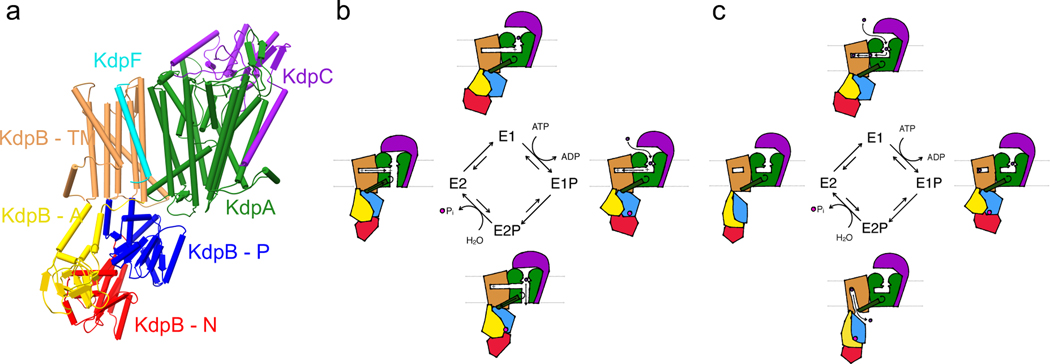
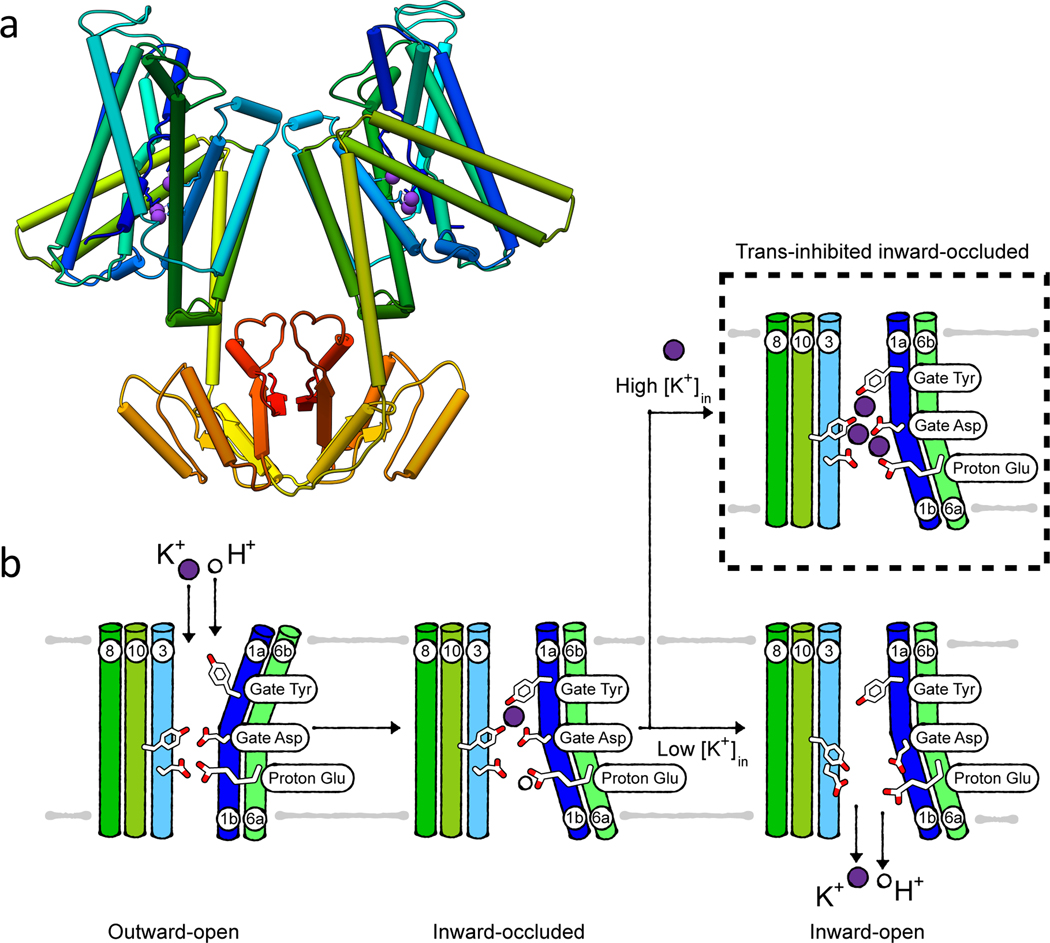
References
-
- Benarroch JM, Asally M. The Microbiologist’s Guide to Membrane Potential Dynamics. Trends Microbiol. 2020;28:304–14. - PubMed
-
- Zhou M, Morais-Cabral JH, Mann S, MacKinnon R. Potassium channel receptor site for the inactivation gate and quaternary amine inhibitors. Nature. 2001;411:657–61. - PubMed
-
- Vieira-Pires RS, Szollosi A, Morais-Cabral JH. The structure of the KtrAB potassium transporter. Nature. 2013;496:323–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
