

The Novel Genetic Background of Infectious Bursal Disease Virus Strains Emerging from the Action of Positive Selection
- PMID: 33801413
- PMCID: PMC7998436
- DOI: 10.3390/v13030396
The Novel Genetic Background of Infectious Bursal Disease Virus Strains Emerging from the Action of Positive Selection
Abstract
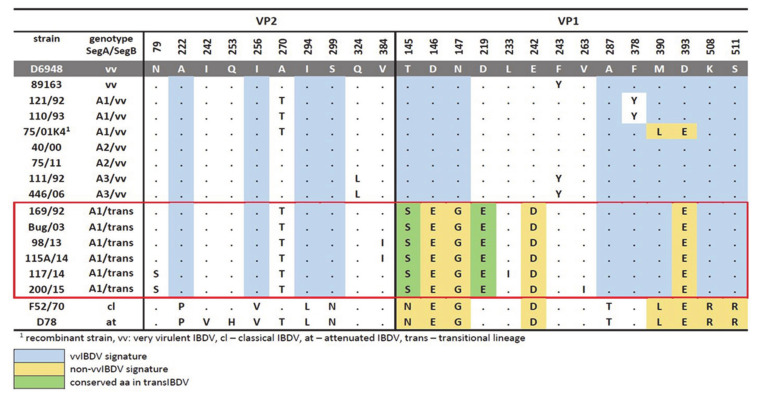
The circulation in Europe of novel reassortant strains of infectious bursal disease virus (IBDV), containing a unique genetic background composition, represents a serious problem for animal health. Since the emergence of this novel IBDV mosaic was first described in Poland, this scenario has become particularly attractive to uncover the evolutionary forces driving the genetic diversity of IBDV populations. This study additionally addressed the phenotypic characterization of these emergent strains, as well as the main features affecting the viral fitness during the competition process of IBDV lineages in the field. Our results showed how different evolutionary mechanisms modulate the genetic diversity of co-existent IBDV lineages, leading to the error catastrophe effect, Muller ratchet effect, or prevalence, depending on their genetic compositions. We also determined that the action of the positive selection pressure, depending on the genomic segment on which it is acting, can drive two main phenotypes for IBDV: immune-escaping strains from the selection on segment A or strains with functional advantages from the selection on segment B. This last group seems to possess an increased fitness landscape in the viral quasispecies composition, presenting better adaptability to dissimilar environmental conditions and likely becoming the dominant population. The reassortant strains also exhibited a lower mortality rate compared with the well-known vvIBDV strains, which can facilitate their spreading.
Keywords: genetic evolution; in vivo pathogenicity; reassortment; very virulent infectious bursal disease virus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Infectious bursal disease virus in Algeria: Detection of highly pathogenic reassortant viruses.Infect Genet Evol. 2018 Jun;60:48-57. doi: 10.1016/j.meegid.2018.01.029. Epub 2018 Feb 1. Infect Genet Evol. 2018. PMID: 29409800
-
Pathogenicity of Genome Reassortant Infectious Bursal Disease Viruses in Chickens and Turkeys.Avian Dis. 2016 Dec;60(4):765-772. doi: 10.1637/11409-031116-Reg. Avian Dis. 2016. PMID: 27902905
-
Identification and Pathogenicity Evaluation of a Novel Reassortant Infectious Bursal Disease Virus (Genotype A2dB3).Viruses. 2021 Aug 25;13(9):1682. doi: 10.3390/v13091682. Viruses. 2021. PMID: 34578267 Free PMC article.
-
[Naturally occurring reassortants of infectious bursal disease virus - A review].Wei Sheng Wu Xue Bao. 2016 May 4;56(5):740-6. Wei Sheng Wu Xue Bao. 2016. PMID: 29727135 Review. Chinese.
-
Genetic Insight into the Interaction of IBDV with Host-A Clue to the Development of Novel IBDV Vaccines.Int J Mol Sci. 2023 May 4;24(9):8255. doi: 10.3390/ijms24098255. Int J Mol Sci. 2023. PMID: 37175960 Free PMC article. Review.
Cited by
-
Purifying selection decreases the potential for Bangui orthobunyavirus outbreaks in humans.Virus Evol. 2023 Mar 8;9(1):vead018. doi: 10.1093/ve/vead018. eCollection 2023. Virus Evol. 2023. PMID: 37025159 Free PMC article.
-
First Detection and Molecular Characterization of Novel Variant Infectious Bursal Disease Virus (Genotype A2dB1b) in Egypt.Viruses. 2023 Dec 7;15(12):2388. doi: 10.3390/v15122388. Viruses. 2023. PMID: 38140629 Free PMC article.
-
The early SARS-CoV-2 epidemic in Senegal was driven by the local emergence of B.1.416 and the introduction of B.1.1.420 from Europe.Virus Evol. 2022 Mar 21;8(1):veac025. doi: 10.1093/ve/veac025. eCollection 2022. Virus Evol. 2022. PMID: 35371561 Free PMC article.
-
Genetics and Pathogenicity of Natural Reassortant of Infectious Bursal Disease Virus Emerging in Latvia.Pathogens. 2022 Sep 22;11(10):1081. doi: 10.3390/pathogens11101081. Pathogens. 2022. PMID: 36297138 Free PMC article.
-
Infectious Bursal Disease Virus in Algeria: Persistent Circulation of Very Virulent Strains in Spite of Control Efforts.Animals (Basel). 2024 Dec 8;14(23):3543. doi: 10.3390/ani14233543. Animals (Basel). 2024. PMID: 39682508 Free PMC article.
References
-
- Cosgrove A.S. An apparently new disease of chickens: Avian nephrosis. Avian. Dis. 1962;6:385–389. doi: 10.2307/1587909. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
