

Ferlins and TgDOC2 in Toxoplasma Microneme, Rhoptry and Dense Granule Secretion
- PMID: 33803212
- PMCID: PMC7999867
- DOI: 10.3390/life11030217
Ferlins and TgDOC2 in Toxoplasma Microneme, Rhoptry and Dense Granule Secretion
Abstract
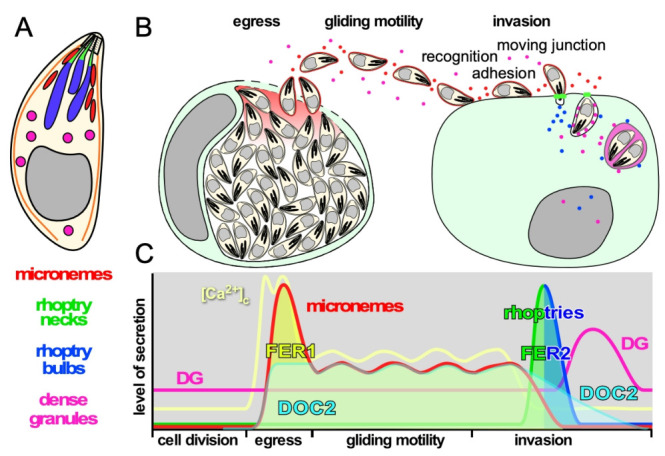
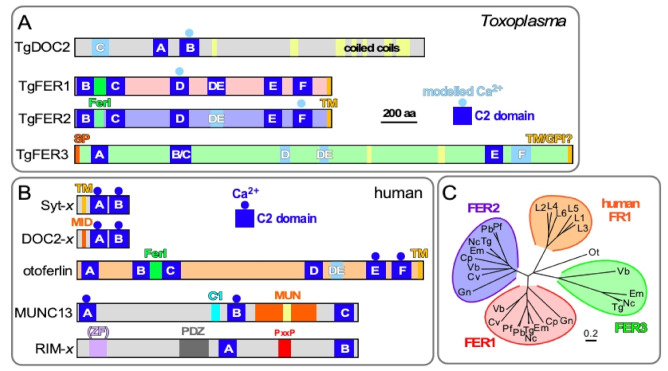
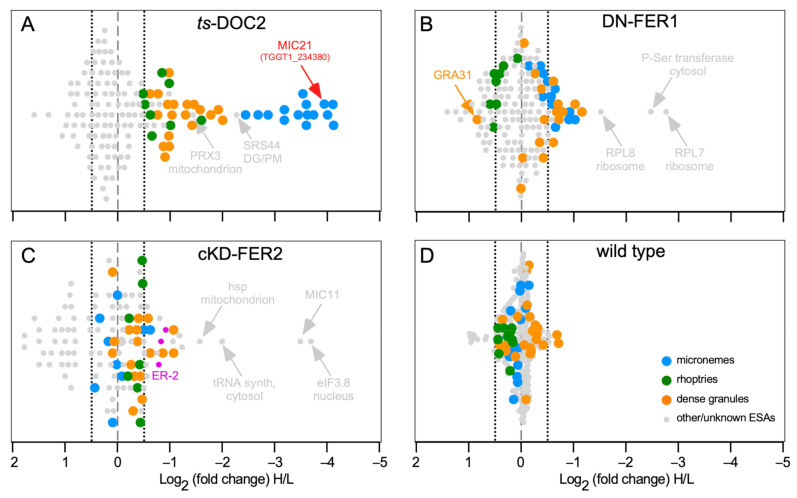
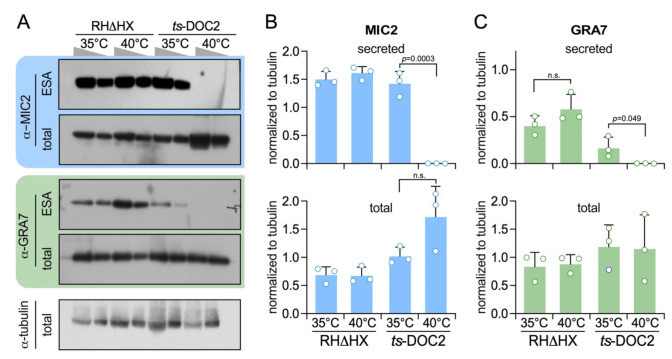
The host cell invasion process of apicomplexan parasites like Toxoplasma gondii is facilitated by sequential exocytosis of the microneme, rhoptry and dense granule organelles. Exocytosis is facilitated by a double C2 domain (DOC2) protein family. This class of C2 domains is derived from an ancestral calcium (Ca2+) binding archetype, although this feature is optional in extant C2 domains. DOC2 domains provide combinatorial power to the C2 domain, which is further enhanced in ferlins that harbor 5-7 C2 domains. Ca2+ conditionally engages the C2 domain with lipids, membranes, and/or proteins to facilitating vesicular trafficking and membrane fusion. The widely conserved T. gondii ferlins 1 (FER1) and 2 (FER2) are responsible for microneme and rhoptry exocytosis, respectively, whereas an unconventional TgDOC2 is essential for microneme exocytosis. The general role of ferlins in endolysosmal pathways is consistent with the repurposed apicomplexan endosomal pathways in lineage specific secretory organelles. Ferlins can facilitate membrane fusion without SNAREs, again pertinent to the Apicomplexa. How temporal raises in Ca2+ combined with spatiotemporally available membrane lipids and post-translational modifications mesh to facilitate sequential exocytosis events is discussed. In addition, new data on cross-talk between secretion events together with the identification of a new microneme protein, MIC21, is presented.
Keywords: Apicomplexa; DOC2; Toxoplasma; dense granule; exocytosis; microneme; rhoptry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Toxoplasma FER1 is a versatile and dynamic mediator of differential microneme trafficking and microneme exocytosis.Sci Rep. 2024 Sep 18;14(1):21819. doi: 10.1038/s41598-024-72628-0. Sci Rep. 2024. PMID: 39294204 Free PMC article.
-
A Member of the Ferlin Calcium Sensor Family Is Essential for Toxoplasma gondii Rhoptry Secretion.mBio. 2018 Oct 2;9(5):e01510-18. doi: 10.1128/mBio.01510-18. mBio. 2018. PMID: 30279285 Free PMC article.
-
A conserved complex of microneme proteins mediates rhoptry discharge in Toxoplasma.EMBO J. 2023 Dec 1;42(23):e113155. doi: 10.15252/embj.2022113155. Epub 2023 Oct 27. EMBO J. 2023. PMID: 37886905 Free PMC article.
-
How Apicomplexa Parasites Secrete and Build Their Invasion Machinery.Annu Rev Microbiol. 2022 Sep 8;76:619-640. doi: 10.1146/annurev-micro-041320-021425. Epub 2022 Jun 7. Annu Rev Microbiol. 2022. PMID: 35671531 Review.
-
The calcium signaling toolkit of the Apicomplexan parasites Toxoplasma gondii and Plasmodium spp.Cell Calcium. 2015 Mar;57(3):186-93. doi: 10.1016/j.ceca.2014.12.010. Epub 2014 Dec 31. Cell Calcium. 2015. PMID: 25605521 Free PMC article. Review.
Cited by
-
Toxoplasma bradyzoites exhibit physiological plasticity of calcium and energy stores controlling motility and egress.Elife. 2021 Dec 3;10:e73011. doi: 10.7554/eLife.73011. Elife. 2021. PMID: 34860156 Free PMC article.
-
Toxoplasma gondii phosphatidylserine flippase complex ATP2B-CDC50.4 critically participates in microneme exocytosis.PLoS Pathog. 2022 Mar 24;18(3):e1010438. doi: 10.1371/journal.ppat.1010438. eCollection 2022 Mar. PLoS Pathog. 2022. PMID: 35325010 Free PMC article.
-
Toxoplasma gondii C2 Domain Protein Deletion Mutant as a Promising Vaccine Against Toxoplasmosis in Mice.Microb Biotechnol. 2025 May;18(5):e70143. doi: 10.1111/1751-7915.70143. Microb Biotechnol. 2025. PMID: 40407413 Free PMC article.
-
Neospora caninum as delivery vehicle for anti-PD-L1 scFv-Fc: A novel approach for cancer immunotherapy.Mol Ther Oncol. 2025 Mar 19;33(2):200968. doi: 10.1016/j.omton.2025.200968. eCollection 2025 Jun 18. Mol Ther Oncol. 2025. PMID: 40236994 Free PMC article.
-
An unconventional SNARE complex mediates exocytosis at the plasma membrane and vesicular fusion at the apical annuli in Toxoplasma gondii.PLoS Pathog. 2023 Mar 27;19(3):e1011288. doi: 10.1371/journal.ppat.1011288. eCollection 2023 Mar. PLoS Pathog. 2023. PMID: 36972314 Free PMC article.
References
-
- Carruthers V.B., Sibley L.D. Sequential protein secretion from three distinct organelles of toxoplasma gondii accompanies invasion of human fibroblasts. Eur. J. Cell Biol. 1997;73:114–123. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
